

Review Article
Volume-1 Issue-1, 2025
Enzyme Properties of Milk-Clotting Enzyme from A Newly Isolated Bacillus Licheniformis D3.11
-
Received Date: March 04, 2025
-
Accepted Date: March 22, 2025
-
Published Date: March 29, 2025
Journal Information
Abstract
In this study, a bacterium producing extracellular milkclotting enzyme was isolated from the soil in Tibetan Plateau, which was identified as Bacillus licheniformis D3.11 using 16S rDNA. Then the milk-clotting enzyme from B. licheniformis D3.11 was purified to 26.25-fold with 18.39% recovery by precipitation in ammonium sulfate and ion-exchange chromatography. The molecular mass of the enzyme was 56.1 kDa as determined by SDS-PAGE, and the result of pepstatin A inhibition treatment showed that this enzyme was an aspartic proteas. The enzyme was active in the pH range 5.5–9.5 and was inactivated completely by heating at 60°C for 20 min. The highest level of enzyme activity was obtained at 60°C, pH 5.5. The high level of milk-clotting activity coupled with a low level of thermal stability suggested that the milk-clotting enzyme from licheniformis D3.11 was a potential substitute for calf rennet.
Key words
Milk-clotting Enzyme; Bacillus licheniformis; Enzyme Properties
Strain |
Milk-clotting activity (SU/mL) |
Proteolysis activity (U/mL) |
Strain |
Milk-clotting activity (SU/mL) |
Proteolysis activity (U/mL) |
D3.11 |
1215.6±3.0 |
14.6±0.5 |
HB4 |
51.5±2.2 |
17.5±1.3 |
HN2 |
21.1±4.2 |
38.3±0.8 |
HX1 |
62.4±3.5 |
36.6±2.3 |
HN3 |
11.3±1.9 |
59.5±2.8 |
HX2 |
116.5±0.9 |
31.9±2.5 |
HN4 |
26.4±3.1 |
46.6±3.1 |
HX3 |
39.5±2.2 |
15.5±1.3 |
GL1 |
16.1±0.6 |
21.9±1.2 |
HX4 |
311.6±5.0 |
14.8±0.6 |
GL2 |
81.2±2.1 |
17.5±2.1 |
HM1 |
26.7±4.2 |
28.3±0.7 |
GL3 |
111.6±5.0 |
14.6±0.6 |
HM2 |
78.3±1.1 |
39.5±2.1 |
GL4 |
126.7±4.2 |
28.3±0.7 |
HM3 |
72.4±3.5 |
36.6±1.0 |
HB1 |
18.3±1.9 |
39.5±2.3 |
HM4 |
26.5±0.9 |
31.9±1.5 |
HB2 |
22.4±3.5 |
36.6±1.1 |
HM5 |
59.5±2.2 |
17.5±1.3 |
HB3 |
16.5±0.9 |
31.9±1.5 |
|
|
|
Purification steps |
Milk- clotting activity (SU) |
Protein content(mg/ ml) |
Specific activity(SU/ mg) |
Purification fold |
Recovery (%) |
Crude enzyme |
1215.6 |
0.315 |
3859.05 |
- |
100.00 |
(NH4)2SO4 |
16021.3 |
0.821 |
12206.21 |
3.16 |
49.25 |
DEAE-SephadexA-25 |
721.5 |
0.0042 |
100357.14 |
26.25 |
18.39 |
Inhibitor |
Concentration (mM) |
Residual activity (%) |
Control |
0 |
100±0.23 |
EDTA |
10 |
94.38±0.93 |
Iodoacetamide |
2 |
95.76±0.93 |
PMSF |
10 |
96.99±1.0 |
Pepstatin A |
0.02 |
1.95±0.15 |
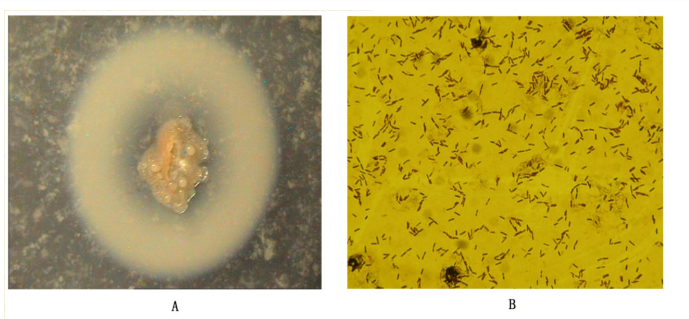 |
| Figure 1: Gram stainand electron micrograph of strain HN1(A); Colony morphology of strain D3.11(B) |
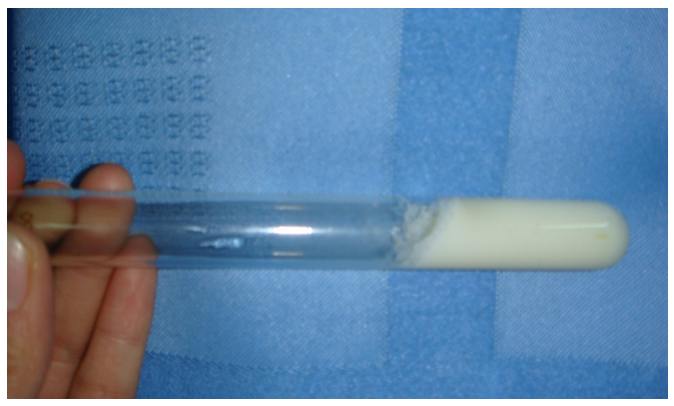 |
| Figure 2:Coagulation of skimmed milk by crude enzyme from D3.11 |
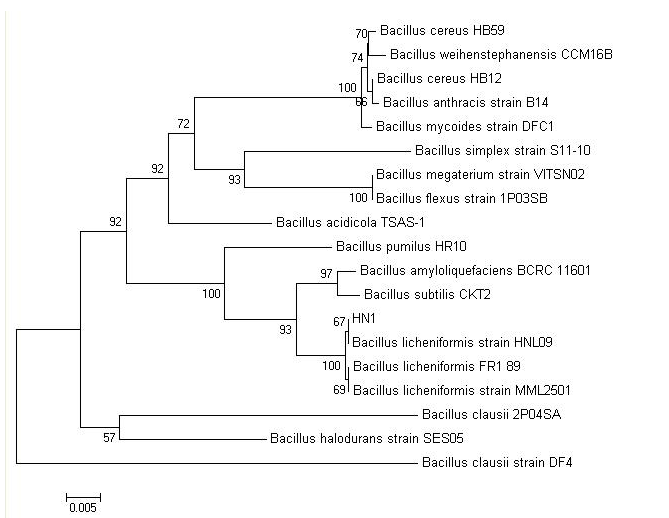 |
| Figure 3: Phylogenetic tree based on 16S rDNA sequence of homology |
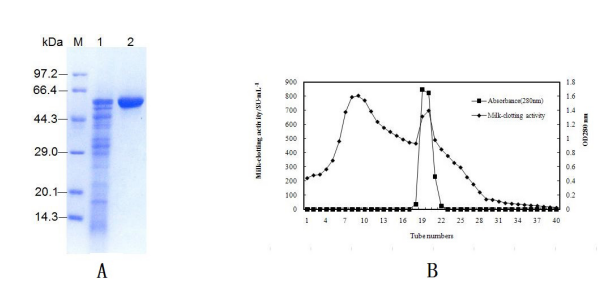 |
| Figure 4: Ion-exchange chromatography through DEAE-Sephadex A-25 of MCE(A); SDS-PAGE electrophoretogram of MCE after purification. Marker lane, standard molecular mass markers; lane 1, MCE purified by precipitation in (NH4)2SO4; lane 2, MCE purified by ion-exchange chromatography through DEAE-Sephadex A-25(B) |
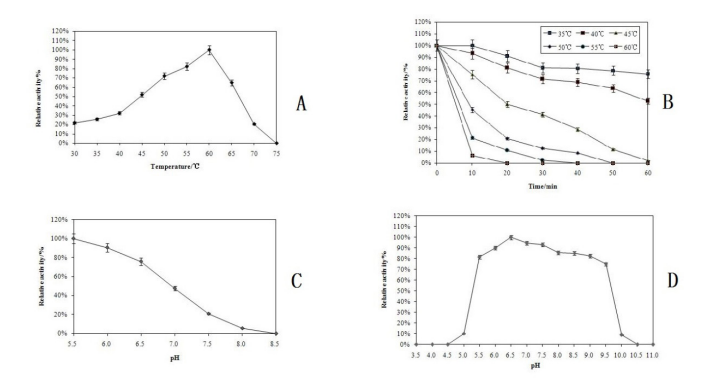 |
| Figure 5: Effect of temperature on milk-clotting activity(A); Thermo-stability of milk-clotting enzyme(B); Influence of different pH on enzyme activity(C); pH stability of milk-clotting enzyme(D) |
Conclusion
Calf rennet, the traditional milk-clotting enzyme, has been widely used by the dairy industry for the manufacture of cheese with good flavour and texture [1, 2]. The worldwide increase of cheese production coupled with a reduced supply of calf rennet has prompted a search for calf rennet substitutes, including milkclotting enzyme from microbe and plant [3]. However, most plant rennets have proved unsuitable because they impart a bitter taste to the cheese [4]. Attention has been focused on the milkclotting enzymes from microbial sources because of its extensive sources, lower cost, and easy extraction [2].
Beside various fungi, numerous bacteria belonging to Bacillus have been suggested as promising microbial rennet producers, such as Bacillus subtilis [5], Bacillus amyloliquefaciens [6], and Bacillus polymyxa [7]. Bacterial rennet appears to be more promising because its production is cheaper, its biochemical diversity is greater, and its genetic modification is easier [6]. However, to obtain the enzyme with high milk-clotting activity, many researchers focus on isolating new bacteria producing milk-clotting enzyme from special environment.
The Tibetan Plateau in China is home to a diversity of microbial species and plentiful microbial germplasm resources [8, 9]. Qinghai pastoral area located in the northwest of the Tibetan Plateau in China. Local herdsmen produce a variety of dairy products such as yoghourt, cheese, Qula and butter from yak milk for thousands of years [10]. It is possible to find microorganisms producing milk-clotting enzyme in this region. The objectives of this study were to screen and isolate microorganisms producing milk-clotting enzyme using wheat bran as nutrient source in this region, and characterize the milk-clotting enzyme produced by B. licheniformis D3.11.
Materials and Methods
Isolation of bacteria producing milk-clotting enzyme
The soil samples were collected from the 5-10 cm layers below soil surface of the yak herding regions in Qinghai pastoral area of China. All soil samples were kept at 4℃ and immediately taken back to the laboratory [6].
One gram of soil was suspended with 9 mL of saline water and a series of ten-fold dilutions of the suspension was made by pipetting 1 mL aliquots into 9 mL saline water. 0.1 mL of each dilution of the series was cultivated at 37°C for 2 days in casein agar plates as the procedure described in [11].
Identification of the selected strain
The selected strain with highest milk-clotting activity was identified using 16S rDNA. Homology searches were performed against the sequences with the database using the BLAST program (NCBI). For phylogenetic analysis, a dataset containing GenBank 16S rRNA gene sequences were aligned using the CLUSTALX program [12]. To construct the phylogenetic tree, MEGA 4.0 software was used [13].
Production of milk-clotting enzyme
Prior to the cultivation of the isolate on the basal fermentation medium, it was cultivated on a seed culture medium (g/L) containing peptone 10, beef extract 3, NaCl 5, with pH 7.2. Incubation was carried out at 37oC, in a rotary shaker, with stirring at 170 rpm for 24 h. The seed cultivation medium was then inoculated aseptically at 2% (v/v) of the basal fermentation medium, and incubated at 37oC, in a rotary shaker, with stirring at 170 rpm for 2 days. The basal fermentation medium contained glucose 5 g, Na2HPO4 0.2 g, with pH 6.2 (w⁄100mL wheat bran juice; wheat bran juice: 10 g wheat bran in 100 mL distilled water, boiled 10 min and filtered through gauze).
Enzyme purification
The crude enzyme was harvested by centrifugation at 8000g for 10 min and then precipitated with ammonium sulfate (30%– 80% saturation). The precipitate obtained after centrifugation at 12,000g for 15 min and suspended in 50 mM sodium phosphate buffer (pH 7.0) and dialysed (7 kDa cutoff) overnight against several changes of distilled water to remove the salt.
Ten millilitres of the partially purified enzyme solution were subjected to ion-exchange chromatography on a DEAESephadex A-25 column (30 cm × 2.6 cm) pre-equilibrated with 50 mM sodium phosphate buffer (pH 7.0). The column was eluted at a flow rate of 0.8 mL/min with an increasing linear gradient of NaCl from 0 to 0.5 M in 50 mM sodium phosphate buffer (pH 7.0) and 5 mL fractions were collected. The protein content of each fraction was determined by measuring the absorbance at 280 nm. The fractions with enzyme activity were pooled and dialysed overnight against distilled water and then lyophilized.
Effects of temperature on enzyme activity
The optimum temperature for the activity of the enzyme was determined by assaying the milk-clotting activity at intervals of 5℃ in the temperature ranging from 30℃ to 75℃.
Effects of temperature on enzyme stability
To determine the thermal stability, the enzyme was incubated at 5℃ intervals ranging from 35℃ to 60℃, and the length of the incubation was varied from 0 to 60 min. After incubation, the residual milk-clotting activity was determined and the activity obtained with an incubation time of 0 min was taken to be 100%.
Effects of pH on enzyme activity
The optimum pH for the activity of the enzyme was determined by assaying the milk-clotting activity in the pH range 5.5–8.5, by adjusting the pH of the substrate (skim milk) with 0.1 M HCl or 0.1 M NaOH as appropriate. The maximum milk-clotting activity obtained was taken to be 100%.
Effects of pH on enzyme stability
To determine the pH stability, the enzyme was dispersed (1:1, v/v) in the following 0.1 M buffer solutions: glycine–HCl (pH 3.5–4.0), citrate/phosphate (pH 4.5–5.5), sodium phosphate (pH 6.0–8.5) and carbonate/bicarbonate (pH 9.0–11.0), and kept at room temperature for 24 h. The residual milk-clotting activity was determined and the maximum activity obtained was taken to be 100%.
Effect of inhibitors
Different protease inhibitors, including a serine-protease inhibitor (phenylmethylsulfonyl fluoride (PMSF) at 10 mM), a metalloprotease inhibitor (ethylene-diaminetetraacetic acid (EDTA) at 10 mM), an aspartic protease inhibitor (pepstatin A at 20 μM) and a cysteine-protease inhibitor (iodoacetamide at 2 mM ) were added separately to the purified enzyme. The mixture was incubated at room temperature for 30 min and the residual milk-clotting activity was tested. The milk-clotting activity obtained without the inhibitors was taken to be 100%.
Milk-clotting activity determination
The milk-clotting activity was determined as described Arima etal [14]. A 5 mL portion of the substrate (10% skim milk in 10 mM CaCl2) was incubated for 5 min at 35oC and then 0.5 mL of enzyme extract was added. The length of time starting from the addition of the enzyme extract to the formation of the first particles was recorded, and the milk-clotting activity was calculated. One Soxhlet Unit (SU) of milk-clotting activity was defined as the amount of enzyme required to clot 1 mL of substrate within 40 min at 35℃.
Proteolysis activity determination
The proteolytic activity of the enzyme was assayed after Arima et al [14]. Enzyme extract (0.5 mL) was added to 2.5 mL of 1% (w/v) alkali soluble casein in 20 mM potassium phosphate buffer (pH 6.5). The mixture was incubated at 35oC in a water bath for 10 min and the reaction was terminated by adding 2.5 mL of 0.44 M trichloroacetic acid, then the mixture was filtered through Whatman No.1 filter paper. One milliliter of 1 M Folin-phenol reagent and 2.5 mL of 0.55 M sodium carbonate solutions was added to 1 mL of the filtrate. This was further incubated for 20 min at 35oC for colour development; optical density (OD) at 660 nm was measured. One unit (U) of enzyme activity was taken as the amount of enzyme, which liberated 1μg of tyrosine per mL per minute.
Results and Discussion
Isolation of bacterium producing milk-clotting enzyme
Table 1 shows the milk-clotting activity and proteolysis activity of different strains isolated from the samples. A total of 22 isolates were obtained for milk-clotting enzyme production, milk-clotting activity of which range from 16.5 to 1215.6 SU/mL. Among them, an isolate D3.11 have the highest milk-clotting activity (1215.6 SU/mL) and lowest proteolysis activity (14.6 U/ mL), which is a rod-shaped and Gran-positive bacterial strain (Figure 1A, Figure 1B). As shown in Fig.2, fermentation broth of D3.11 greatly coagulated the skim milk.
The 16S rDNA gene sequences of D3.11 were compared to all sequences in GenBank and showed the closest match to that of Bacillus licheniformis HNL09, Bacillus licheniformis FR189 and Bacillus licheniformis MML2501 with a homology of 99%. For phylogenetic analysis, the phylogenetic tree (Figure 3) was constructed using neighbor-joining. Therefore, the isolate D3.11 could be identified as Bacillus licheniformis. Bacillus licheniformis D3.11 has been deposited in the China General Microbiological Culture Collection Center with CGMCC NO.3289.
Purification of the enzyme
Because the purity of an enzyme can impact the product quality, the milk-clotting enzyme of B.licheniformis D3.11 was purified by ammonium sulphate precipitation and column chromatography with DEAE-Sephadex A-25. The results of the purification procedure are summarized in Table 2. Partial purification of the enzyme, 3.16-fold purification and 49.25% recovery was achieved by precipitation in 30–80% saturated ammonium sulfate. Passage through a DEAE-Sephadex A-25 column eluted with a gradient of NaCl further purified the enzyme to 26.25-fold with 18.39% recovery. The enzyme has a molecular mass of 56.1 kDa as determined by SDS-PAGE (Fig.4A). The elution pattern of the milk-clotting enzyme after ion-exchange chromatography (DEAE-Sephadex A-25 column, elution with a linear gradient of 0–0.5 M NaCl) is shown in Figure 4B.
Effects of temperature on enzyme activity and stability
Milk-clotting enzymes of different origins have different optimum temperatures. As was showed in Fig 5A, the milkclotting activity increased with increased temperature in the temperature range 30–55°C and the optimum temperature for the purified enzyme was 60°C. The heat stability of the purified enzyme is shown in Figure 5B. It was fully active after 10 min of incubation at 35°C and retained 75.7% activity after 60 min. At 40°C, the enzyme showed good stability and retained 81.1% activity after 20 min. But above 50°C, it caused greatly loss in activity. As soon as the temperature was up to 60°C, the milkclotting activity was completely lost.
Effects of pH on enzyme activity and stability
The maximum milk-clotting activity was at pH 5.5 for the purified enzyme, and the activity decreased with increasing pH (Figure 5C). The stability of the purified enzyme at different pH values is shown in Figure 5D. The enzyme was stable in a relatively wide range of pH 5.5–9.5, with maximum stability at pH 6.5. Outside either end of this range, the activity of the enzyme decreased drastically, which showed that the B. licheniformis D3.11 enzyme was stable in the acidic to neutral range.
Effect of inhibitors
In order to determine the type of milk-clotting protease produced by D3.11, inhibition experiments with different protease inhibitors were conducted. Table 3 showed the sensitivity of the purified enzyme to a serine protease inhibitor (PMSF), a cysteine protease inhibitor (iodoacetamide), a metalloprotease inhibitor (EDTA) and an aspartic protease inhibitor (pepstatin A). The fact that PMSF, EDTA and iodoacetamide did not inhibit the enzyme activity showed that the enzyme was not a serine protease, metalloprotease or cysteine protease. The strong inhibition, 97.83% at 5 mM pepstatin A, showed that the enzyme belongs to the aspartic protease group.
Discussions
Many bacteria especially several species belonging to Bacillus are known to produce variety of extracellular enzymes, such as amylases, protease, and xylanase [15, 16]. Numerous bacteria such as B. amyloliquefaciens, B. subtilis and B. subtilis natto have been suggested as promising microbial rennet producers [17, 18, 19, 20]. In the present study, several bacteria producing milk-clotting enzyme isolated from soil samples collecting from Qinghai pastoral area in Tibetan Plateau of China. Among these bacteria, B. licheniformis D3.11 possessed high rennetproducing capacity. To our knowledge, this is the first report on B. licheniformis producing milk-clotting enzymes that was isolated from Qinghai pastoral area in Tibetan Plateau of China.
Milk-clotting enzymes from different sources have different enzyme properties. The enzyme from B. licheniformis D3.11 has a molecular mass of 56.1 kDa, which is close to milk-clotting enzymes from B. amyloliquefaciens D4 [20] and higher than others (34–49 kDa) [21, 22, 23, 24, 25]. The optimum temperature of the enzyme from B. licheniformis D3.11 was different from calf rennet, which has an optimum temperature in the range 40–42°C. Compared with the milk-clotting enzymes from other microorganism such as B. sphaericus [2], Mucor pusillus [24] and Rhizomucor miehei [22] the B. licheniformis D3.11 enzyme has a lower level of thermostability. Similar to calf rennet, the purified enzyme from B. licheniformis D3.11 had a higher level of milkclotting activity in the acidic range. The low pH-sensitivity and thermostability of the enzyme is useful for cheese making.
Wheat bran, an agro-industrial residue, contains cellulose material, starch, crude protein, trace elements and other certain ingredients. Wheat bran is abundant and cheap, usually used as feedstuff. There are several reports describing wheat bran as potent substrate for enzyme production [20]. In the present study, B. licheniformis D3.11 possessed high rennet-producing capacity. Moreover, the medium used for enzyme production is wheat bran. These results suggest that B. licheniformis D3.11 is a possible commercial source of milk-clotting enzyme for cheese making [26].
Conclusion
In this study, a total of 22 isolates were obtained for milk-clotting enzyme production and an isolate with good milk-clotting activity identified as Bacillus licheniformis and the properties of the milk-clotting enzyme were investigated. The purified enzyme from B. licheniformis D3.11 belongs to the aspartic protease group with a molecular mass of 56.1 kDa, and has low thermostability and low sensitivity to pH. The result suggest that B. licheniformis D3.11 is a possible commercial source of milk-clotting enzyme for cheese-making. An evaluation of its potential for use in cheese-making will be the subject of future work.
Acknowledgements
The authors are grateful to the National Natural Science Fund of China (31760466), and the Natural Science Fund of Gansu Province (20JR10RA524) for financial support of this work.
References
- McSweeney PLH and Fox PF (1997) Chemical methods for the characterization of proteolysis in cheese during ripening. Lait 77: 41-76.
- Magda AE (2004) Formation and properties of serine protease enzyme with milk-clotting activity from Bacillus sphaericus. Egypt J. Appl. Sci. 19: 68-91.
- Cavalcanti MTH, Teixeira MFS, Filho JLL, and Porto ALF (2004) Partial purification of new milk-clotting enzyme produced by Nocardiopsis sp. Bioresource Technol. 93: 29-35.
- Raposo S and Domingos A (2008) Purification and characterization milk-clotting aspartic proteinases from Centaurea calcitrapa cell suspension cultures. Process Biochem. 43: 139-44.
- Dutt K, Gupta P and Saran S (2009) Production of milkclotting protease from Bacillus subtilis. Appl Biochem Biotech 158: 761-72.
- He XL, Zhang WB, Ren FZ, Song X, Gan BZ, et al. (2012) Screening fermentation parameters of the milk-clotting enzyme produced by newly isolated Bacillus amyloliquefaciens D4 from the Tibetan Plateau in China. Annals of Microbiology, 62: 357-65.
- Magda A, El-Bendary, ME Moharam and TH Ali (2007) Purification and Characterization of Milk Clotting Enzyme Produced by Bacillus sphaericus. J Appl Sci Res. 3: 695-9.
- Zhang SH, Liu XH, Liang F, Wang L, Liu YH, et al. (2009) Screening of cryophilic cellulose-degradable bacterium and its zymological properties. J Microbiol 29: 97-100.
- Wang GX, Huang HH, and Zhang M (2010) Isolation and identification of a fibrinolytic bacterium strain from frozen soil in the Tibetan Plateau. Acta Microbiological Sinica. 50: 148-54.
- Liu HN, Zhang C, Zhang H, Guo HY, Wang PJ, et al. (2013) pH treatment as an effective tool to select the functional and structural properties of yak milk caseins. Journal of Dairy Science. 96: 5494-500.
- Arima K, Iwasaki S, and Tamura G (1967) Milk clotting enzyme from microorganisms. Part I: screening test and identification of the potent fungus. J. Agr. Biol. Chem. 31:540-5.
- Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F and Higgins DG (1997) The CLUSTALX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25: 4876-82.
- Tamura K, Dudley J, Nei M, and Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol 24: 1596-9.
- Arima K, Yu J, and Iwasaki S (1970) Milk-clotting enzyme from Mucor pusillus var. Lindt. Method. Enzymol. 19:446-59.
- Deng AH, Wu J, Zhang Y, Zhang GQ and Wen TY (2010) Purification and characterization of a surfactant-stable highalkaline protease from Bacillus sp. B001. Bioresource Technol 18 :7100-6.
- Bindu B, Saurabh SD, Sonia A, Ritu M and Jitender S (2012) Application of thermostable xylanase of Bacillus pumilus in Textile Processing. Indian J Microbiol 52:222-9.
- Ageitos JM, Vallejo JA and Sestelo ABF (2007) Purification and characterization of a milk-clotting protease from Bacillus licheniformis strain USC13.J Appl Microbiol 103: 2205-13.
- Shieh CJ, Phan TL and Shih IL (2009) Milk-clotting enzymes produced by culture of Bacillus subtilis natto. Biochem Eng J 43: 85-91.
- Ding ZY, Liu SP, Gu ZH, Zhang L, Zhang KC, et al. (2011) Production of milk-clotting enzyme by Bacillus subtilis B1 from wheat bran. Afr J Biotechnol 10: 9370-8.
- Zhang WB, He XL, Liu HN, Ren FZ, Guo HY, et al. (2013) Statistical optimization of culture condition for milk-clotting enzyme production by Bacillus amyloliquefaciens using wheat bran-an agro-industry waste. Indian J Microbiol 54: 492-5.
- Sardinas JL (1968) Rennin Enzyme of Endothia parasitica. Appl. Microbiol. 16: 248-55.
- Preetha S and Boopathy R (1997) Purification and characterization of a milk clotting protease from Rhizomucor miehei. World J. Microb. Biot. 13: 573-8.
- Kumar S, Sharma NS, Saharan MR, and Singh R (2005) Extracellular acid protease from Rhizopus oryzae: Purification and characterization. Process Biochem 40: 1701-5.
- Nouani A, Belhamiche N, Slamani R, Belbraouet S, Fazouane F, et al. (2009) Extracellular protease from Mucor pusillus: purification and characterization. Int. J. Dairy Technol. 62: 112-7.
- Vishwanatha KS, Rao AGA, and Singh SA (2010) Production and characterization of a milk-clotting enzyme from Aspergillus oryzae MTCC 5341. Appl. Microbiol. Biot. 85: 1849-59.
- Das K, Doley R and Mukherjee AK (2004) Purification and biochemical characterization of a thermostable, alkaliphilic, extra-cellular a-amylase from Bacillus subtilis DM-03, a strain isolated from the traditional fermented food of India. Appl Biochem Biotech 40: 291-8.
Artcle Information
Review Article
Received Date: March 04, 2025
Accepted Date: March 22, 2025
Published Date: March 29, 2025
Journal of Foodscience Nutrition and Public Health
Volume 1 | Issue 1
Citation
Zhongming Zhana, Haijun Qiao, Weibing Zhang (2025) Enzyme Properties of Milk-Clotting Enzyme from A Newly Isolated Bacillus Licheniformis D3.11. J Food Sci Nutr Public Health 1(1):102
Copyright
©2025 Weibing Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

doi: jese.2025.1.102