

Review Article
Volume-1 Issue-1, 2025
Effect of Azide on Haematococcus pluvialis
Received Date: March 05, 2025
Accepted Date: March 23, 2025
Published Date: March 30, 2025
Journal Information
Abstract
Haematococcus pluvialis can produce and accumulate large amounts of astaxanthin under stress conditions. In this study we investigated influence of different concentrations of azide (0, 25, 50 and 100 µM), in two-time periods (pretreatment- 7 days and treatment- 14 days) to compare the changes on growth and biochemical parameters in green and red stages. Azide treatment and preptreatment caused a decrease in all measured parameters except of carotenoid in green stage. Carotenoid content did not show any changes in azide treatment but pre-treatment with low concentration induced carotenoid accumulation in green stage. The amount of dry weight, protein and carbohydrate did not change in red cells that had treated with azide, but carotenoid content decreased in these cells. Pre-treated with azide had a negative effect on the amount of protein and carbohydrate but increased the carotenoid content. Azide pre-treatment had better performance in increasing the amount of carotenoids in red cell. These results show that H. pluvialis offer potential to phytoremediation of azide and carotenoid accumulation.
Key words
Sodium Azide; Carbohydrate; Carotenoid;Protein; Pretreatment
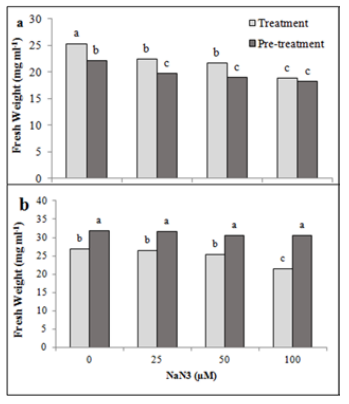 |
| Figure 1: Effect of azide treatment and pre-treatment on fresh weight in the green (a) and red (b) stage of H. pluvialis. Values are means of three-replications ±standard deviation. Different letters show the significant difference with p< 0.05 in one-way ANOVA and Duncan's tests |
 |
| Figure 2: Effect of azide treatment and pre-treatment on dry weight in the green (a) and red (b) stage of H. pluvialis. Values are means of three-replications ±standard deviation. Different letters show the significant difference with p< 0.05 in one-way ANOVA and Duncan's tests |
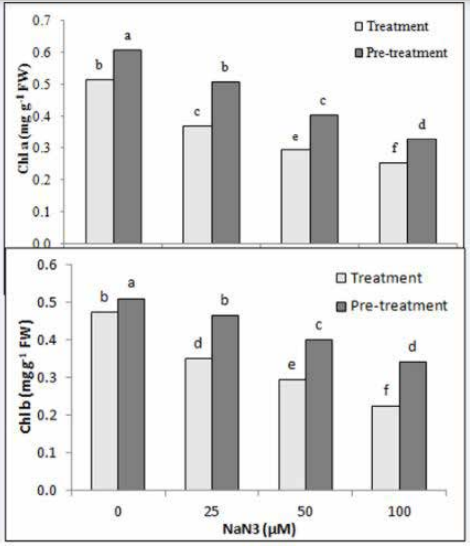 |
| Figure 3: Effect of azide treatment and pre-treatment on chlorophyll content in the green stage of H. pluvialis. Values are means of three-replications ±standard deviation. Different letters show the significant difference with p< 0.05 in one-way ANOVA and Duncan's tests |
 |
| Figure 4: Influence of Methanolic Extractives of Prepupal Stages of Black Soldier Fly (MEPSBSF) on Activity of Asparateaminotransferase (AST) Alanine aminotransferase (ALT) and Lactate Dehydrogenase (LDH) in Serum in DMBA Induced Toxicity in Norwegian Rat, Rattus norvegicus (L) |
 |
| Figure 5: Effect of azide treatment and pre-treatment on carbohydrate content in the green (a) and (b) red stage of H. pluvialis. Values are means of three-replications ±standard deviation. Different letters show the significant difference with p< 0.05 in one-way ANOVA and Duncan's tests |
 |
| Figure 6: Effect of azide treatment and pre-treatment on protein content in the green (a) and (b) red stage of H. pluvialis. Values are means of three-replications ±standard deviation. Different letters show the significant difference with p< 0.05 in one-way ANOVA and Duncan's tests |
Introduction
The Chlorophyte alga H. pluvialis (Volvocales) is usually found in temperate regions around the world (Czygan 1970; Lorenz 1999). Haematococcus has a rather complex life cycle with two distinct cell forms, green flagellated motile cells and non-motile non-flagellated cysts formed under stress conditions. Stress effects on astaxanthin production in Haematococcus pluvialis under different culture conditions (Sarada et al. 2002). The green vegetative cells contain chlorophyll and small amount of carotenoids. Algae cells accumulate carotenoids under stress conditions such as nitrogen deficiency, high radiation or other tension processes. This stage is usually accompanied by the transformation of green vegetative cells into red cysts. At first astaxanthin is accumulated in lipid droplets in the cytoplasm surrounding the nucleus, then massive accumulation of carotenoids occurs and astaxanthin-containing lipid droplets eventually fill the whole cytoplasm. Among commercial microalgae, H. Pluvialis is the richest astaxanthin sources, which has been shown to be a potent antioxidant and significantly scavenges free radicals and reduces oxidative stress (O'Connor and O'Brien 1998). As a strong inhibitor of lipid peroxidation it plays an active role in protecting biological membranes from oxidative damage (Palozza and Krinsky 1992). The algae also contains significant amounts of protein, fat, and carbohydrates (Goodenough and Heuser 1985). Different factors such as light, culture medium compositions, pH, growth regulators, and temperature are effective to achieve maximum biomass and astaxanthin production. These factors usually have opposite effect on astaxanthin content and biomass production. It means stress conditions that increase astaxanthin content, on the other hand, decrease growth and biomass production. Sometimes, if the cells are exposed to severe stress, growth is stopped, and the cells die at a relatively short time (Su et al. 2014).
Different methods such as genetic and metabolic engineering, and synthetic biology can be used for manipulation of biological systems especially microbes for different purposes. However, genetic manipulation leads to permanent changes in a species that might have adverse effects when used for environmental and agricultural applications (Hunt et al. 2010). Improvement of microalga biomass and bioactive compounds productivity using alternative means such as chemicals can be a safe method in this regards.
Studies have indicated that biochemical stimulants offer significant potential to increase microalgae productivity (Cheng et al. 2012). It has been shown that increasing of reactive oxygen species (ROS) induce astaxanthin accumulation in H. pluvialis under normal growth conditions and it is possible that the carotenogenic response is mediated by ROS (Kobayashi et al. 1993). Azide is an inhibitor of the enzyme cytochrome c oxidase in the fourth complex of the electron transport chain. The disruption of the electron transport chain causes ROS produced and induction of oxidative stress (Apel and Hirt 2004). In microorganisms, alternative oxidase (AOX) can serve as a terminal electron acceptor in the presence of inhibitors of the cytochrome c oxidase electron transport chain (ETC), such as azide and cyanide (Trumpower 1990). It was showed that AOX activity have important influence in plant growth. The amount of AOX protein was effective to maintain photosynthetic performance and improved growth during prolonged water deficit in N. tabacum (Dahal and Vanlerberghe 2018).
The present study investigated the effect of azide as a respiratory inhibitor in two-time period (7 and 14 days) on growth and biochemical parameters of H. pluvialis.
Methods and Materials
Haematococcus pluvialis (UTEX) grown in autotrophic Bold’s Basal medium (Tripathi et al. 1999). The cultures were incubated at 25±1 °C under 16/8 h light/dark cycle with 50 µmol m2 s1 intensity. The initial cell concentration was adjusted to 3.5 x104 cells ml-1. The cultures were grown in 250 ml Erlenmeyer flasks with 150 ml of medium under two different conditions: 1) pretreatment, the algae were cultured in the medium with different concentrations of sodium azide (0, 25, 50 and 100 μM) for 7 days, and then were transferred to a fresh medium without sodium azide for a further 7 days, 2) treatment, algae were exposed to different concentrations of sodium azide (0, 25, 50 and 100 μM) for 14 days. Then all cultures (treatment and pre-treatment) were harvested by centrifugation at 3500 g for 5 min and resuspended in fresh BG11 medium without nitrogen (Stanier et al. 1971) to induce red cells formation for 14 days.
Growth was measured by counting cell numbers using a haemacytometer. Dry weight of the algal biomass was estimated after drying at 60 °C in a hot-air oven until a constant weight was obtained. For pigment analysis, a known quantity of biomass was extracted with 96% methanol and, chlorophyll and carotenoids were quantified as per the procedure given by Şükran et al, (Şükran et al. 1998).
Soluble protein concentration was obtained according to Bradford (Bradford 1976). To assay soluble carbohydrate content Fales (Fales 1951) method was used.
All experiments were performed in three replicate. Data are presented as mean ± standard deviation (SD), and they were analyzed by one-way analysis of variance, and the significance of the differences (P < 0.05) was estimated by Duncan’s multiple comparison test.
Results and Discussion
Algae treated and pretreated with azide showed fresh weight loss of green cells (Figure 1a). However, these treatments did not have a significant effect on the fresh weight of red cells except for cells treated with 100 μM azide which showed less fresh weight compared to control cells (Figure 1b). Azide treatment significantly decreased dry weight of algae cells (Figure 2). This effect was dose dependent. The lowest dry weight was observed in culture treated by 100 µM azide with 6-time decrease. Pre-treatment by azide also declined cell dry weight of H. pluvialis in green phase, but it was not dose dependent. There was not significant difference between applied concentrations of azide. Dry weight in the treated algae was about 60% control. An amazing result was that azide treatment and pre-treatment in green stage did not have effect on dry weight in red stage, just 100 µM azide treatment caused a 28% decrease in dry weight in compared to control cells. Our results showed that the azide concentrations used in this study had toxic effects on H. pluvialis physiology. Azide is a known metabolic inhibitor in plants and algae with many potential targets. Azide binds irreversibly to the heme cofactor in cytochrome C oxidase, inhibits catalase and superoxide dismutase, which scavenge ROS, therefore, it enhances oxidative stress (Zalogin and Pick 2014). Indeed, reduction in growth and biochemical parameters can be due to oxidative stress created by azide. It was reported that hydrogen peroxide production was enhanced several fold in Anabaena nidulans by treatment with azide (Morales et al. 1992). The addition of azide had no significant effect on cell biomass of Anabaena and Dunaliella tertiolecta (Nultsch et al. 1983; Chen et al. 2019). Contrary to our results, an experiment performed on different algae, reported that azide-treated cells had higher rate of photosynthesis, more chlorophyll, and faster growth (Zalogin and Pick 2014).
Both treated and pre-treated algae with azide had lower amount of chlorophyll a and b than control cells (Figure 2). The lowest amount of chlorophyll a was observed in 100 µM azide treatment and pre-treatment with 50% control cultures. The amount of chlorophyll b showed a 53 and 33% decrease in comparison to control in treatment and pretreatment by 100 µM azide, respectively. Similar conclusions have been reported in Dunaliella tertiolecta and Eruca sativa and it was indicated that azide had a certain inhibitory effect on the accumulation of chlorophylls and carotenoids in algae cells and the inhibitory effect was proportional to the concentrations of azide (Chen et al. 2019).
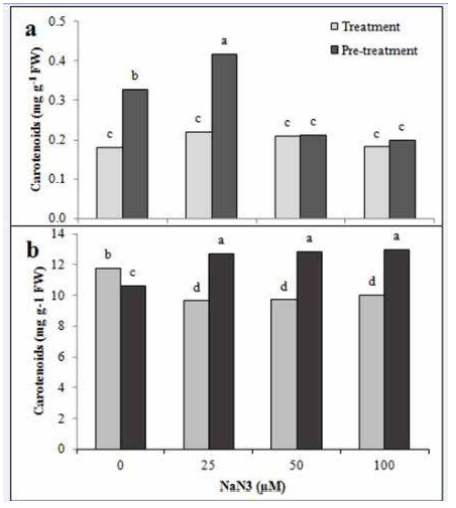
Azide treatment had no significant effect on the carotenoid content of green cells (Figure 3). Pre-treatment of azide at 25 μM concentration resulted in a significant increase in carotenoids (28% increase compared to control). There was no significant difference between 50 and 100 μM concentrations of azide pretreatment, but these two concentrations significantly decreased the carotenoid level compared to the control. A decrease in the carotenoid content of D. tertiolecta and Eruca sativa was reported in cultures treated with azide (Chen et al. 2019; AlQurainy 2009).
The results of the effect of treatment and pre-treatment of azide on the carbohydrate content of green cells are shown in Figure 4. No significant difference was observed in carbohydrate content between the control and 25 and 50 μM azide treatments, although the amount of carbohydrate significantly decreased at 100 μM azide. In the pre-treatment of sodium azide, the amount of carbohydrate in control and concentration of 25, 50 and 100 μM azide were 3.13, 2.35, 1.75 and 1.42 mg g-1 FW, respectively. By increasing azide concentration, carbohydrate content reduced to half of carbohydrate content in control (by 100 μM azide).
Adding azide to culture medium in green phase for 14 days did not affect carbohydrate content of cells in red phase (Figure 4). Azide Pre-treatment of cells in green phase significantly reduced carbohydrate content in red cells. The effects of azide on the reduction of carbohydrate content was dose dependent. The lowest amount of carbohydrate was observed in 100 μM azide with almost 50% control. Carbohydrate reduction can be the result of depletion of chlorophyll and photosynthesis rate. Inhibitory effect of azide on photosynthesis was proved in plants (Forti and Gerola 1977).
The results showed that the amount of protein of green cells decreased in azide treatment and pre-treatment (Figure 5). The lowest protein content was observed in algae treated and pre-treated by 100 μM azide with 54 and 50% control respectively. The results of measuring the protein content of red cells H. pluvialis under sodium azide treatment showed that sodium azide treatment did not have a significant effect on the protein content of red cells. Sodium azide pre-treatment reduced the protein content of red cells significantly. The highest amount of protein was observed in pre-treatment red cells in the control group and the lowest amount of protein was observed in pre-treatment red cells at 100 μM concentration (50% reduction compared to control). As mentioned previously inhibitory effects of azide on nitratereductase was confirmed (Zalogin and Pick 2014). This enzyme has key role in nitrogen assimilation; therefore, reducing the amount of protein in the presence of azide is a predictable result (Figure 6).
Conclusion
Azide with the concentrations used in this experiment showed negative effects on the growth and biochemical parameters of the algae. Only the carotenoid content of green cells increased by 27% with 25 μM azide pretreatment and red by 18% with red pretreatment of all azide concentrations.
Acknowledgment
This work was carried out at Shahid Bahonar University of Kerman, Iran.
References
- Al-Qurainy F (2009) Effects of sodium azide on growth and yield traits of Eruca sativa (L.). World Applied Sciences Journal 7: 220-6.
- Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55: 373-99.
- Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry 72: 248-54.
- Chen HH, Xue LL, Liang MH, Jiang JG (2019) Sodium azide intervention, salinity stress and two-step cultivation of Dunaliella tertiolecta for lipid accumulation. Enzyme and microbial technology 127:1-5.
- Cheng JS, Niu YH, Lu SH, Yuan YJ (2012) Metabolome analysis reveals ethanolamine as potential marker for improving lipid accumulation of model photosynthetic organisms. J Chemical Technology & Biotechnology 87: 1409-18.
- Czygan F (1970) Blood-rain and blood-snow: nitrogen-deficient cells of Haematococcus pluvialis and Chlamydomonas nivalis. Archiv fur Mikrobiologie 74: 69-76.
- Dahal K, Vanlerberghe GC (2018) Improved chloroplast energy balance during water deficit enhances plant growth: more crop per drop. J experimental botany 69: 1183-97.
- Fales F (1951) The assimilation and degradation of carbohydrates by yeast cells. Journal of Biological Chemistry 193: 113-24
- Forti G, Gerola P (1977) Inhibition of photosynthesis by azide and cyanide and the role of oxygen in photosynthesis. Plant physiology 59: 859-62.
- Goodenough UW, Heuser JE (1985) The Chlamydomonas cell wall and its constituent glycoproteins analyzed by the quickfreeze, deep-etch technique. The Journal of cell biology 101: 1550-68.
- Hunt RW, Chinnasamy S, Bhatnagar A, Das K (2010) Effect of biochemical stimulants on biomass productivity and metabolite content of the microalga, Chlorella sorokiniana. Applied Biochemistry and Biotechnology 162: 2400-14.
- Kobayashi M, Kakizono T, Nagai S (1993) Enhanced carotenoid biosynthesis by oxidative stress in acetate-induced cyst cells of a green unicellular alga, Haematococcus pluvialis. Applied and Environmental Microbiology 59: 867-873
- Lorenz RT (1999) A technical review of Haematococcus algae. NatuRoseTM Technical Bulletin 60:1-12
- Morales I, Batuecas S, de la Rosa FF (1992) Storage of solar energy by production of hydrogen peroxide by the blue‐green alga Anacystis nidulans R2: Stimulation by azide. Biotechnology and bioengineering 40: 147-50.
- Nultsch W, Schuchart H, Koenig F (1983) Effects of sodium azide on phototaxis of the blue-green alga Anabaena variabilis and consequences to the two-photoreceptor systems-hypothesis. Archives of microbiology 134: 33-7.
- O'Connor I, O'Brien N (1998) Modulation of UVA light-induced oxidative stress by β-carotene, lutein and astaxanthin in cultured fibroblasts. J Dermatological Science 16: 226-30.
- Palozza P, Krinsky NI (1992) Astaxanthin and canthaxanthin are potent antioxidants in a membrane model. Archives of biochemistry and biophysics 297: 291-5.
- Sarada R, Tripathi U, Ravishankar G (2002) Influence of stress on astaxanthin production in Haematococcus pluvialis grown under different culture conditions. Process Biochemistry 37: 623-7.
- Stanier R, Kunisawa R, Mandel M, Cohen-Bazire G (1971) Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriological reviews 35: 171.
- Su Y, Wang J, Shi M, Niu X, Yu X, Gao L, Zhang X, Chen L, et al. (2014) Metabolomic and network analysis of astaxanthin-producing Haematococcus pluvialis under various stress conditions. Bioresource technology 170: 522-9.
- Şükran D, GÜNEŞ T, Sivaci R (1998) Spectrophotometric determination of chlorophyll-A, B and total carotenoid contents of some algae species using different solvents. Turkish Journal of Botany 22: 13-8.
- Tripathi U, Sarada R, Rao SR, Ravishankar G (1999) Production of astaxanthin in Haematococcus pluvialis cultured in various media. Bioresource Technology 68: 197-9.
- Trumpower BL (1990) Cytochrome bc1 complexes of microorganisms. Microbiology and Molecular Biology Reviews 54: 101-29.
- Zalogin TR, Pick U (2014) Inhibition of nitrate reductase by azide in microalgae results in triglycerides accumulation. Algal Research 3: 17-23.
Artcle Information
Review Article
Received Date: March 05, 2025
Accepted Date: March 23, 2025
Published Date: March 30, 2025
Journal of Microbiology and Bacteriology Research
Volume 1 | Issue 1
Citation
Hakimeh Mansouri (2025) Effect of Azide on Haematococcus pluvialis. J Microbiol and Bacteriol Res 1:103
Copyright
©2025 Hakimeh Mansouri. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

doi: jmbr.2025.1.103