

Review Article
Volume-1 Issue-1, 2025
Therapeutic Potential of NRF2 pathway in Neurological Disorder
-
Received Date: June 05, 2025
-
Accepted Date: June 23, 2025
-
Published Date: June 30, 2025
Journal Information
Abstract
Oxidative stress play a major role in the cause of many diseases including neurological disorders such as Alzheimer's, and Parkinson's to maintain homeostasis between free radicles and antioxidants. certain pathways protect the nerve cell and release antioxidants to reduces oxidative stress. Nrf2 activation has a promising therapeutic effect on many diseases including neuroprotection by releasing more than 250cytoprotective genes and also known as the master regulatory pathway of Antioxidants. SIRTUIN 3 activation releases Antioxidants genes SOD2, catalase to reduces ROS and increase mitochondrial biogenesis. AMPK activation promotes autophagy by ULK1 and activatesPGC-1alpha which is the master regulator of mitochondrial biogenesis. Heat shock protein activation promotes chaperonmediated autophagy and macroautophagy to perform neuroprotection against accumulated protein.
Key words
Neurodegeneration, Oxidative Stress, Nrf2, Heat Shock Protein
Abbreviations :NRF2: (Nuclear erythroid factor or Nuclear respiratory factor); KEAP-1(Kelch-like ECH associating protein); BTB domain (Broad complex tram track & Bric a brack); IVR (The intervening region); DGR (Double glycine repeats); (bzip) basic leucine zipper CNC; maf (masculoaponeurotic fibrosarcoma oncogene); ARE (Antioxidative response elements); (UBA) ubiquitin association; GSK3(Glycogen synthase Kinase 3); SOD2(superoxide dismutase enzyme ); OPA1 Optic atrophy 1; PINK 1(phosphatase and tensin homolog PTEN, induced kinase); NMNAT2( Nicotinamide mononucleotide adenyltransferase); NAD( Nicotinamide adenine dinucleotide); 3-TYP(Triazol pyridine); Trimethylamine-N-Oxide (TMAO), 5AMPK Activation ( Adenosine monophosphate-activated protein kinase), 00=(HMGCR) 3-Hydroxy 3-methylglutary Co-A reductase; TSC2(Tumur suspension gene); mTOR(mamalin target of rapamycin); PGC-1alpha (Peroxisome proliferator activator receptor gamma coactivator 1- alpha); CPT-1 (carnitine palmitoyltransferase-1); RabGAP (RabGTPase activating protein); ERR(Estrogen related receptor); (OMPs)outer membrane proteins HSP (Heat shock protein)
Company/Compound |
Comments |
Ref |
Reata /Bordoxolone Methyl |
synthetic triterpenoid is in PhaseIII It shows pulmonary aterial hypertension .In phase II it shows chronic kidney failure and II diabetics |
54 |
Reata/omaveloxolone |
second-generation synthetic (2019) triterpenoid is in Phase for |
|
Biogen: ALK8700 |
It is an prodrug of monomethylfumarte for multiple sclerosis is in Phase |
|
Evgen Pharma: SFX-01 |
Cyclodextrin+Sulforaphane are in phase II for subarachnoid hemorrrhage ; effective and safe In metastatic breast cancer with |
|
Anthocyanin |
It is an flavonoid present In a garpes and berries used in atherosclerosis |
|
ITH12674 |
Hybrid molecule of melatonin+sulforaphane for brain ischemia by modify the Cys-151 of keap for NRF2 activation is in preclincalstudies |
TITLE |
REFE |
Nrf2-ARE pathway: An emerging target against oxidative stress and neuroinflammation in |
37 |
Nrf2-Keap1 signaling in oxidative and reductive stress |
38 |
The Nrf2/ARE pathway: A promising target to counteract mitochondrial dysfunction in Parkinson’s |
39 |
The Nrf2-ARE pathway: A valuable therapeutic target for the treatment of neurodegenerative diseases. |
40 |
The cytoprotective role of the Keap1-Nrf2 pathway. |
41 |
Sirtuin deacetylases in neurodegenerative diseases of aging. |
42 |
SIRT3 Regulation of Mitochondrial Quality Control in Neurodegenerative Diseases |
43 |
The role of SIRT3 in the brain under physiological and pathological conditions. |
44 |
Mitochondrial Sirtuin 3: New emerging biological function and therapeutic target. |
45 |
The AMPK signalling pathway coordinates cell growth, autophagy and metabolism |
46 |
The crosstalk between Nrf2 and AMPK signal pathways is important for the anti-inflammatory effect of Berberine in LPS-stimulated macrophages and endotoxin-shocked mice. Antioxidants Redox Signal |
47 |
Orientin-mediated Nrf2/HO-1 signal alleviates H2O2-induced oxidative damage via induction of JNK |
48 |
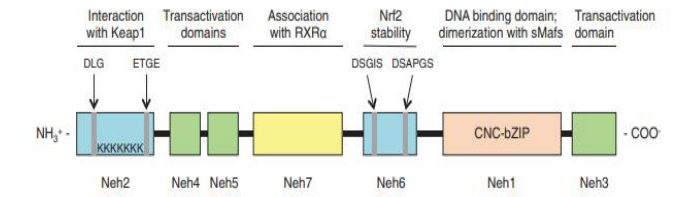 |
| Figure 1: human Nrf2 protein structure. The Nrf2 protein is made up of seven Neh domains. The Neh1 CNC-bZIP domain is in charge of DNA binding and dimerization with small Maf proteins; the Neh2 domain mediates interaction with Keap1 via the DLG and ETGE motifs and contains seven lysine residues that are ubiquitylated; the Neh3, Neh4, and Neh5 domains are transactivation domains; the Neh6 domain is a serine-rich region that regulates Nrf2 stability; and the Neh7 domain bZIP, basic-region leucine zipper; CNC, cap ‘‘n" collar; Keap1, Kelch-like ECHassociated protein 1; Neh, Nrf2-ECH homology; Nrf2, nuclear factor E2-related factor 2; RXRa [4]. |
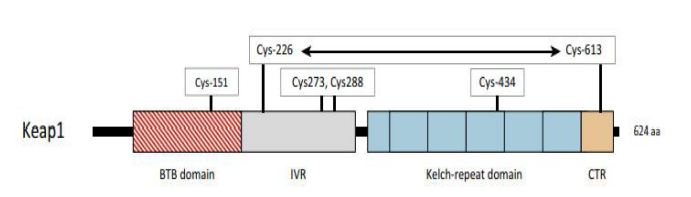 |
| Figure 2:The domains of Keap1 are displayed in their current locations. The Bric-a`-Brac (BTB) domain (shown as a striped red box) is responsible for the formation of Keap1 homodimers and the recruitment of Cullin-3, whereas the Kelch-repeat domain (each Kelch domain is depicted as a blue box) and the C-terminal region (CTR, depicted in light orange) form a six-bladed β-propeller structure that binds the DLG and ETGE motifs in Nrf2. The intervening region (IVR), which wraps around the β-propeller structure, is made up of residues between the BTB and Kelch-repeat domains. Residues that function as sensors for electrophiles and H2O2 (i.e., Cys-151, Cys-226/Cys-613, Cys-273/Cys-288, and Cys-434) are shown in open boxes above the cartoon, and the horizontal two-headed arrow between Cys-226 and Cys-613 signifies that they form a disulfide bridge when exposed to H2O2 [10]. |
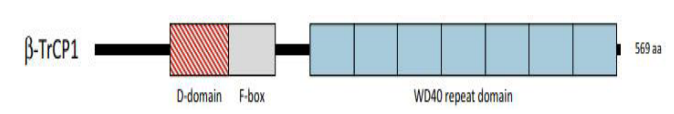 |
| Figure 3:The domain locations in b-TrCP1 are shown. A striped red box represents the dimerization domain (D domain), which produces homo- and heterodimers between b-TrCP1 and b-TrCP2, and a grey box represents the F-box, which recruits Skp1 (S-Phase Kinase-Associated Protein 1). Finally, the blue boxes indicate the WD40 repeat domain, which binds the DSGIS and DSAPGS motifs in Nrf2 [10]. |
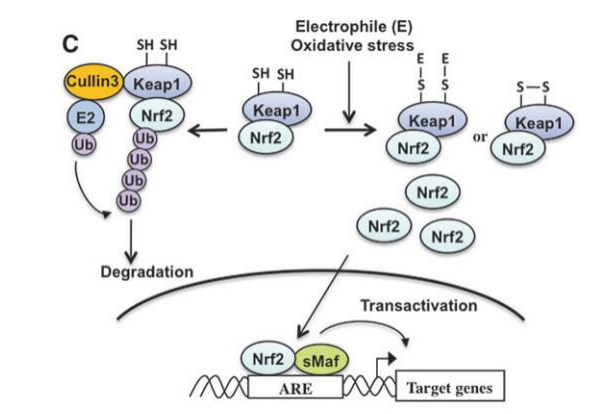 |
| Figure 4: The mechanism of Nrf2 activation is depicted in this diagram. Nrf2 is constitutively ubiquitinated by the Keap1=Cullin3 ubiquitin ligase complex in non-stressed circumstances. Keap1 is inactivated and changed at its regulatory cysteine residues when exposed to electrophile (E) or oxidative stress. Inactivation of Keap1 promotes nuclear accumulation of Nrf2 and ARE-dependent transcription. [15]. |
Introduction
Parkinson's disease (PD) is a neurodegenerative condition characterised by the gradual loss of midbrain dopaminergic neurons, as well as mitochondrial failure, and is characterised by the cardinal symptoms of rigidity, bradykinesia, tremor, and postural instability [11]. Neurological diseases are currently the major cause of disability across the world. PD was the fastest increasing neurological illness in terms of prevalence, disability, and mortality in the Global Burden of Disease (GBD) research. According to recent studies, the global number of persons affected by the condition more than quadrupled between 1990 and 2016 [12].
Nuclear factor erythroid 2-related factor 2 (Nrf2)
In both glial cells and neurons, NRF2 is an essential defender against injuries in the brain. As well as influencing several antioxidant enzymes, NRF2, Phase I and II drug metabolising enzymes and mitochondrial pathways can also enhance the expression of anti-inflammatory mediators [17].
Nrf2 knock-out mice, cortical culture has demonstrated they are susceptible to oxidative stress, Nrf2 re-expression increases neuroprotection through ARE activation [18].
Nrf2 Expression in Neurodegenerative Diseases
According to a research done by Kaimin Chan et al., Nrf2 is expressed at low levels in many tissues, at high levels in the liver, lungs, and kidneys, and at the greatest levels in the cell lining of the digestive system, which extends from the oesophagus to the small and large intestines. In the CNS, Nrf2 is expressed in the lateral ventricle and its medial wall, as well as the choroid plexus in the fourth ventricle. Nrf2 expression is also found in the olfactory epithelium in the nasal cavity, the thyroid and submandibular glands, and the brown fat layer in the back. At higher magnification, NRF2 expression is most pronounced in the luminal cells of the stomach and intestine [19].
In Alzheimer's disease, Nrf2 is primarily expressed in cytoplasmic hippocampus neurons; it does not translocate to the nucleus; a biochemical extract of the frontal cortex confirmed decreased levels of nuclear Nrf2 in Alzheimer's disease. In the nuclei of Alzheimer's disease patients, neither neurons nor astrocytes showed substantial Nrf2 staining. The Nrf2 pathway is most likely dysfunctional in AD hippocampal neurons; nevertheless, in Parkinson's disease, Nrf2 is more numerous in dopaminergic neurons than in SNpC (substantia nigra pars compacta), and dopaminergic Nrf2 exhibits neuronal sensitivity to increased oxidative stress [20].
Nrf2 Structure
NRF2 (Nuclear factor erythroid 2-related factor 2) also called Nuclear factor erythroid-derived 2-like 2, was discovered in 1994 laboratory of Yuet Wai Kan, and it belongs cap "n" collar (CNC) subfamily of basic leucine zipper (bZIP) transcription factors and is encoded by NFE2L2 gene [1, 2]. It involves members possessing a homology region of CNC with 43 amino acids, which supports the N-term DNA-binding domain to the DNA-binding specificity of this family [3]. NRF2 is a modular protein that consists of 7 functional domains (Neh 1-7), as seen under Figure 1, known as the NRF2 ECH (Neh) domains, each of which has a distinct function [3, 4]. Neh1 contains a basic leucine zipper motif (bZIP) that heterodimerizes with small musculoaponeurotic fibrosarcoma protein, DNA, and other transcription partners, a study conducted by Plafker et al. Has shown that Neh1 of Nrf2 forms a nuclear complex with ubiquitin-conjugating enzyme UbcM2 utilizing DNA binding and regulating the stability of Nrf2 [5, 6]. Neh2 N-terminus interacts with the kelch domain of KEAP1 (Cullin (Cul)3–RING, dimeric redox-sensitive substrate adaptor Ubiquitin ligase complex including the box protein (Rbx)1 (i.e., CRLKeap1) through two binding sites, the stronger binding ETGE motif and the weaker binding DLG motif [7, 3, 10] shown in Figure 2. Studies show Interaction of C-terminal Neh3 with the transcription coactivator known as CHD6 (a chromo-ATPase/helicase DNAbinding protein) may play a key role in the activation of ARE driven gene [8]. Neh4 and Neh5 bind together to CBP (CREBbinding protein), synergistically stimulate gene expression via ARE and synchronizes reporter gene activation [9]. Neh6 contains two motifs, DSGIS and DSAPGS, which interact with dimeric bTrCP (b-transducin repeat-containing protein), which acts as a substrate receptor for S-phase kinase-associated protein 1 (Skp1)–Cul1– Rbx1 core E3 complex (i.e., SCFb-TrCP) shown in Figure 3, these two motifs are important for Nrf2 stability. It is important to note that Neh6 controls the stability of NRF2 in KEAP1 independent manner, Neh7 suppresses the NRF2–ARE signalling pathway by interacting with the retinoic X receptor alpha (RXR) [5, 10].
Nrf2-KEAP1-ARE pathway
Oxidative stress is one of the major key player in chronic inflammation associated with neurodegeneration, The KEAP1- Nrf2 pathway is a major signaling pathway for antioxidant defense against reactive oxygen species assault. Nrf2 induces expression of antioxidant enzyme genes such as glutathione S-transferase (GST) and NAD(P)H:quinone oxidoreductase 1 (NQO1), as well as hemeoxygenase-1 (HO1). This induction requires antioxidant response element (ARE) DNA sequence [13, 14], KEAP1 suppress activity of Nrf2 by binding to Neh2, it act as adaptor molecule for the Cul3 E3 ligase complex and leads to Nrf2 degradation through the ubiquitin–proteasome pathway, exposure to reactive oxygen species or electrophiles modifies KEAP1 and inactivates there by leading to attenuation of Nrf2 degradation [15], causing accumulation of Nrf2 in the nucleus and heterodimerization with small Maf proteins (MafF, MafG, and MafK) this substance binds to ARE at regulatory region of targeted gene [16,4] shown in Figure 4.
The regulation of Nrf2 is complicated, yet it can be regulated at several levels. Figure 1 depicts the transcriptional and posttranscriptional levels [4]. NRF2 abundance within the cell is tightly regulated and is mainly controlled by four E3 ubiquitin ligase complexes-mediated ubiquitylation and proteasomal degradation: KEAP1-Cullin (CUL) 3-RING-box protein (RBX)1, βTrCP-S-phase kinase-associated protein-1 (SKP1)-CUL1- RBX1, WD Repeat protein (WDR), 3-CUL4-damaged DNA binding protein (DDB) 1, and HRD1 (also called Synoviolin) under different conditions [23].
In unstressed condition its basal(homeostatic) protein level is low reason being KEAP1 mediated proteasomal degradation by Keap1/Cul3 E3 ubiquitin ligase complex [4,23], reactive oxidant species modifys specific cystin residue of KEAP1 (C257, C273, C288, and C297) causing disassociation of KEAP1 and Nrf2 migrate to nucleus causing its accumulation as evidenced in KEAP1 knockdown in human cells and KEAP1 deletion in mice, and activation of cytoprotective genes [22,21,24,25].
Many cytoplasmic protein which have KEAP1 dependent but cysteine independent mechanisms have been identified, that stabilizes Nrf2 by attenuating interaction between KEAP1-Nrf2. P62 also known as Autophagy cargo-adaptor p62/sequestosome 1 (SQSTM1) [26,27,28,29,30,31]. dipeptidyl peptidase 3 (DPP3) [29], Wilms tumor gene on X chromosome (WTX) [32], and Partner and Localizer of BRCA2 (PALB2) [33] all contain KEAP1-interacting region (KIR)-like ETGE motifs and thus competes with NRF2 for KEAP1 binding, resulting in KEAP1 sequestration and NRF2 stabilization. P21 cyclin-dependent kinase p21Cip1/WAF1, interacts with DLG and ETGE inhibits KEAP1-Nrf2 binding [34]. BRCA1 promotes stability and activation of Nrf2 [35], P300 interacts with Nrf2 and interferes with KEAP1-Nrf2 complex formation [36].
NRF2 controls the expression of four genes, which include Glucose 6-phosphate dehydrogenase, 6-phosphogluconate dehydrogenase, Malic enzyme 1, and Isocitrate dehydrogenase 1, which are responsible for the production of NADPH, which acts as a cofactor and fuel for antioxidant reactions. It also controls the expression of cytochrome P450 oxidoreductase. Among blood cells, monocytes and neutrophils have the greatest amounts of NRF2. In astrocytes, NRF2 levels are high. Furthermore, microglia, a monocyte lineage, exhibits higher amounts of NRF2 than neurons [49, 50, 51, 52].
NRF2 lowers inflammation through three different ways, redox metabolism modification, cross-talk with (NF-KB), direct regulation of pro-inflammatory genes [49, 53]. Table 1 gives list of Nrf2 activators.
SITRULIN-Nrf2
SIRT1 is the most extensively researched sirtuin, and it regulates several processes that govern eating and energy expenditure. When it is reduced, insulin resistance increases. If sirtuin1 levels rise, insulin sensitivity rises, sirtuin 1 deacytalates and promotes PGC1 alpha, ERR, which also has cell-protective action in both neuronal and non-neuronal cells by promoting autophagy and controlling cellular cholesterol, sirtuin3 levels rise, and NRF1, NRF2 levels rise. Which demonstrate a defence mechanism against ROS [55,56,57,58].
Heat shock protein (HSP)
Heat shock proteins are a family of proteins produced by stressful conditions by cells. It is generally seen in heat accumulation / stress, cold stress, UV stress, during wound healing and tissue remodeling it activates molecular chaperones to regulate misfolding of new proteins and the negative effect on cellular protein caused by stressors such as high temperature, redox imbalance, heavy metals [59,60,61,62,63].
The upregulation of heat-shock protein is also said to be heat shock response and induced by heat shock factor. This HSP is found in all organisms from bacteria to humans.HSPs classified according to there molecular weight HSP60,HSP70,HSP90. Ubiquitin is a small protein for degradation also comes under HSP [64,65,66]. Activation of HSP in response to environmental stressors has been determined in bacteria. During heat stress, outer membrane proteins (OMPs) get misfolded and accumulated in periplasmic space which is detected by protease which passes the sigma signal transcription factor. Some studies show that accumulation or misfolding of proteins leads to activation of HSP[67].RNA thermometer such as four thermometers, Rose element, and Hsp90 regulatory element is the mechanism for upregulation of heat shock proteins in some bacteria.
Petersen and Mitchell experiment on Drosophila melanogaster to seen the heat shock gene expression in heat stress as well as in cold stress and found that the HSP gene protects from death and enhances survival [68,69,70]. Molecular chaperones are the main function of heat shock protein. They regulate proper protein shape and correction of misfolding proteins by maintaining stabilize unfolded proteins. HSPs help to transfer proteins across the cell. All organism has some members of heat shock proteins family for protein maintenance [71,72]. HSP aids in monitoring proteins in the cell and they carry old proteins or damaged proteins to proteosomes for degradation and prevent new proteins from misfolding this activity are part of the cell repair system also known as a cellular stress response or heat shock response. Heat shock protein shows self-degradation on itself by proteolytic action [73]. HSP84, HSP90, HSP70, HSP27 has significant cardio protective role. HSP90 shows vascular relaxation by binding both soluble guanylate cyclase and endothelial nitric oxide synthase. Gata4 gene regulates cardiac genes hspb12 and hspb7 which are responsible to prevent cardiac myopathies in embryos of zebrafish are observed and show cardiac morphogenesis.HSPb7 also known as cardiac heat shock protein and act on kupffer vesicles for regulation of left-right asymmetry of zebrafish heart.Hsp20 shows a major role in the prevention of platelet aggregation, cardiac myocytes function, skeletal muscle function, muscle insulin response, and prevention of apoptosis after ischemia injury [74,75,76].
Macroautophagy and chaperon-mediated autophagy is promoted by HSPs when damaged protein aggregate and misfolding of synthesized protein [77,78]. Table 2 referenced different pathways for neuroprotective activity.
Conclusion
Neurological disorders such as Parkinson's and Alzheimer's are primarily caused by oxidative stress or redox imbalance, in which free radicals or ROS react with cellular lipids, proteins, and DNA, resulting in cell death. Protein accumulation, mitochondrial dysfunction, proteasomal dysfunction to mitigate these parameters, all of these factors contribute to cell death and neurological disorders. Different detoxification mechanisms can minimise oxidative stress and sustain mitochondrial biogenesis while boosting autophagy for the destruction of accumulated proteins and misfolded proteins.
Conflicts of interest
The authors declare no conflicts of interest concerning authorship or publication of this article
Declaration of competing interest
The authors whose names are listed immediately below certify that they have NO affiliations with or involvement in any organization or entity with any financial interest (such as honoraria; educational grants; participation in speakers’ bureaus; membership, employment, consultancies, stock ownership, or other equity interest; and expert testimony or patent-licensing arrangements), or non-financial interest (such as personal or professional relationships, affiliations, knowledge or beliefs) in the subject matter or materials discussed in this manuscript.
Acknowledgement
The authors would also like to express their special thanks to the management of JSS College of Pharmacy, Ooty and JSS Academy of Higher Education & Research, Mysuru
References
- Moi P, Chan K, Asunis I, Cao A, Kan YW (1994) Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proceedings of the National Academy of Sciences of the United States of America, 91(21), 9926-30.
- Cuadrado A, Rojo AI, Wells G, Hayes JD, Cousin SP, et al. (2019) Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nature reviews. Drug discovery 18: 295-317.
- Song MY, Lee DY, Chun KS, Kim EH (2021) The role of nrf2/ keap1 signaling pathway in cancer metabolism. Int J Mol Sci 22: 1-16.
- Tonelli C, Chio IIC, Tuveson DA (2018) Transcriptional Regulation by Nrf2. Antioxidants and Redox Signaling, 29: 1727-45.
- Akhileshwar Namani, Yulong Li, Xiu Jun Wang, XiuwenTang (2014) Modulation of NRF2 signaling pathway by nuclear receptors: Implications for cancer, Biochim. Biophys. Acta 9: 1875-85.
- Plafker KS, Nguyen L, Barneche M, Mirza S, Crawford D, et al. (2010) The ubiquitin-conjugating enzyme UbcM2 can regulate the stability and activity of the antioxidant transcription factor Nrf2. J of Biol Chem 285: 23064–74.
- Tong KI, Katoh Y, Kusunoki H, Itoh K, Tanaka T, et al. (2006) Keap1 recruits Neh2 through binding to ETGE and DLG motifs: characterization of the two-site molecular recognition model. Mol and Cell Biol 26: 2887-900.
- Nioi P, Nguyen T, Sherratt PJ, Pickett CB (2005) The carboxyterminal Neh3 domain of Nrf2 is required for transcriptional activation. Mol and Cell Biol 25: 10895-906.
- Katoh Y, Itoh K, Yoshida E, Miyagishi M, Fukamizu A, et al. (2001) Two domains of Nrf2 cooperatively bind CBP, a CREB binding protein, and synergistically activate transcription. Genes to cells: devoted to molecular & cellular mechanisms, 6: 857-68.
- Hayes JD, Dinkova-Kostova AT (2014) The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends in biochemical sciences, 39: 199-218.
- Yamaguchi A, Ishikawa K, Tsuyoshi Inoshita, Kahori ShibaFukushima, Shinji Saiki, et al. (2020) Identifying Therapeutic Agents for Amelioration of Mitochondrial Clearance Disorder in Neurons of Familial Parkinson Disease. Stem Cell Rep 14: 1060-75.
- Feigin VL, Vos T (2019) Global, regional, and national burden of neurological disorders, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol 18: 459- 80.
- Kim S, Indu Viswanath AN, Park JH, Lee HE, Park AY, et al. (2020) Nrf2 activator via interference of Nrf2-Keap1 interaction has antioxidant and anti-inflammatory properties in Parkinson's disease animal model. Neuropharmacology 167: 107989.
- Ma Q (2013) Role of nrf2 in oxidative stress and toxicity. Annual review of pharmacology and toxicology, 53: 401-26.
- Itoh K, Mimura J, Yamamoto M (2010) Discovery of the negative regulator of Nrf2, Keap1: a historical overview. Antioxidants & redox signaling, 13: 1665-78.
- Yamazaki H, Tanji K, Wakabayashi K, Matsuura S, Itoh K (2015) Role of the Keap1/Nrf2 pathway in neurodegenerative diseases. Pathology international, 65: 210-9.
- Brandes MS, Gray NE (2020) NRF2 as a Therapeutic Target in Neurodegenerative Diseases. ASN Neuro 12.
- Kraft AD, Johnson DA, Johnson JA (2004) Nuclear factor E2-related factor 2-dependent antioxidant response element activation by tert-butylhydroquinone and sulforaphane occurring preferentially in astrocytes conditions neurons against oxidative insult. J Neurosci: the official journal of the Society for Neuroscience, 24: 1101-12.
- Chan K, Lu R, Chang JC, Kan YW (1996) NRF2, a member of the NFE2 family of transcription factors, is not essential for murine erythropoiesis, growth, and development. Proceedings of the National Academy of Sciences of the United States of America 93: 13943-8.
- Ramsey CP, Glass CA, Montgomery MB, Lindl KA, Ritson GP, et al. (2007) Expression of Nrf2 in neurodegenerative diseases. J of Neuropathol and Exp Neurol 66: 75–85.
- Baird L, Llères D, Swift S, Dinkova-Kostova AT (2013) Regulatory flexibility in the Nrf2-mediated stress response is conferred by conformational cycling of the Keap1-Nrf2 protein complex. Proceedings of the National Academy of Sciences, 110:15259 LP-64.
- Albena T Dinkova-Kostova, W David Holtzclaw, Robert N Cole, Ken Itoh, Nobunao Wakabayashi, et al. (2002) Direct evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidants. Proc Natl Acad Sci USA 99: 11908-13.
- He F, Ru X, Wen T (2020) NRF2, a transcription factor for stress response and beyond. Int J Mol Sci 21: 1-23.
- Wakabayashi N, Itoh K, Wakabayashi J, Motohashi H, Noda S, et al. (2003) Keap1-null mutation leads to postnatal lethality due to constitutive Nrf2 activation. Nature Genetics 35: 238-45.
- Devling TW, Lindsay CD, McLellan LI, McMahon M, Hayes JD (2005) Utility of siRNA against Keap1 as a strategy to stimulate a cancer chemopreventive phenotype. Proceedings of the National Academy of Sciences of the United States of America, 102: 7280-5A.
- Taniguchi K, Yamachika S, He F, Karin M (2016) p62/ SQSTM1-Dr. Jekyll and Mr. Hyde that prevents oxidative stress but promotes liver cancer. FEBS letters, 590: 2375-97.
- Komatsu M, Kurokawa H, Waguri S, Taguchi K, Kobayashi A, et al. (2010) The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nature cell Biol 12: 213-23.
- Umemura A, He F, Taniguchi K, Nakagawa H, Yamachika S, et al. (2016) p62, Upregulated during Preneoplasia, Induces Hepatocellular Carcinogenesis by Maintaining Survival of Stressed HCC-Initiating Cells. Cancer cell 29: 935-48.
- Hast BE, Goldfarb D, Mulvaney KM, Hast MA, Siesser PF, et al. (2013) Proteomic analysis of ubiquitin ligase KEAP1 reveals associated proteins that inhibit NRF2 ubiquitination. Cancer Res 73: 2199-210.
- Kim PK, Hailey DW, Mullen RT, Lippincott-Schwartz J (2008) Ubiquitin signals autophagic degradation of cytosolic proteins and peroxisomes. Proceedings of the National Academy of Sciences of the United States of America 105: 20567-74.
- Lau A, Wang XJ, Zhao F, Villeneuve NF, Wu T, et al. (2010) A noncanonical mechanism of Nrf2 activation by autophagy deficiency: direct interaction between Keap1 and p62. Mol and Cell Biol 30: 3275-85.
- Camp ND, James RG, Dawson DW, Yan F, Davison JM, et al. (2012) Wilms tumor gene on X chromosome (WTX) inhibits degradation of NRF2 protein through competitive binding to KEAP1 protein. J Biol Chem 287: 6539-50.
- Ma J, Cai H, Wu T, Sobhian B, Huo Y, et al. (2012) PALB2 interacts with KEAP1 to promote NRF2 nuclear accumulation and function. Mol and Cell Biol 32: 1506-17.
- Chen W, Sun Z, Wang XJ, Jiang T, Huang Z, et al. (2009) Direct interaction between Nrf2 and p21(Cip1/WAF1) upregulates the Nrf2-mediated antioxidant response. Mol Cell 34: 663-73.
- Gorrini C, Baniasadi PS, Harris IS, Silvester J, Inoue S, et al. (2013) BRCA1 interacts with Nrf2 to regulate antioxidant signaling and cell survival. J Exp Med 210: 1529-44.
- Ganner A, Pfeiffer ZC, Wingendorf L, Kreis S, Klein M, et al. (2020) The acetyltransferase p300 regulates NRF2 stability and localization. Biochem and Biophy Res Comm 524: 895-02.
- Buendia I, Michalska P, Navarro E, Gameiro I, Egea J, et al. (2016) Nrf2–ARE pathway: An emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases . Pharmacology & Therapeutics 157: 84-104.
- Bellezza I, Giambanco I, Minelli A, Donato R (2018) Biochem. Biophys. Acta - Mol Cell Res 1865: 721-33.
- Tufekci KU, Civi Bayin E, Genc S, Genc K (2011) The Nrf2/ARE Pathway: A Promising Target to Counteract Mitochondrial Dysfunction in Parkinson's Disease. Parkinson’s Disease 314082: 14.
- Joshi G, Johnson JA (2012) The Nrf2-ARE Pathway: A Valuable Therapeutic Target for the Treatment of Neurodegenerative Diseases. CNS Drug Discov. 7: 218-29.
- Baird L, Dinkova-Kostova AT, (2011) The cytoprotective role of the Keap1-Nrf2 pathway. Arch Toxicol 85: 241-72.
- Herskovits AZ, Guarente L (2013) Sirtuin deacetylases in neurodegenerative diseases of aging. Cell Res 23: 746-58.
- Meng H, Yan WY, Lei YH, Wan Z, Hou YY, et al. (2019) SIRT3 Regulation of Mitochondrial Quality Control in Neurodegenerative Diseases. Front Aging Neurosci 11: 1-10.
- Sidorova-Darmos E, Sommer R, Eubanks JH (2018) The Role of SIRT3 in the Brain Under Physiological and Pathological Conditions. Front Cell Neurosci 12: 1-16.
- Zhang J, Xiang H, Liu J, Chen Y, He RR, et al. (2020) Mitochondrial Sirtuin 3: New emerging biological function and therapeutic target. Theranostics 10: 8315-42.
- Mihaylova MM, Shaw RJ (2011) The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat Cell Biol 13: 1016-23.
- Mo C, Wang L, Zhang J, Numazawa S, Tang H, et al. (2014) The Crosstalk Between Nrf2 and AMPK Signal Pathways Is Important for the Anti-Inflammatory Effect of Berberine in LPS-Stimulated Macrophages and Endotoxin-Shocked Mice. Antioxidants & Redox Signaling 20: 574-88.
- Xiao Q, Piao R, Wang H, Li C, Song L (2018) Orientinmediated Nrf2/HO-1 signal alleviates H2O2-induced oxidative damage via induction of JNK and PI3K/AKT activation. Int J Biol Macromol 118: 747-55.
- Cuadrado A, Rojo AI, Wells G, Hayes JD, Cousin SP, et al. (2019) Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat Rev Drug Discov 18: 295-317.
- Jong-Min Lee, Marcus J Calkins, Kaimin Chan, Yuet Wai Kan, Jeffrey A Johnson (2003) Identification of the NF-E2- Related Factor-2-Dependent Genes Conferring Protection against Oxidative Stress in Primary Cortical Astrocytes Using Oligonucleotide Microarray Analysis.” J Biol Chem 278: 12029-38.
- Kai Connie Wu, Julia Yue Cui, Curtis D Klaassen (2011) “Beneficial Role of Nrf2 in Regulating NADPH Generation and Consumption.” Toxicol Sci 123: 590-600.
- Mitsuishi, Yoichiro, YukieKawatani, Tatsuhiro Shibata, Toshihiro Nukiwa, et al. (2012) “Nrf2 Redirects Glucose and Glutamine into Anabolic Pathways in Metabolic Reprogramming.” Cancer Cell 22: 66-79.
- Philip Wenzel, Sabine Kossmann, Thomas Münzel, Andreas Daiber, et al. (2017) “Redox Regulation of Cardiovascular Inflammation – Immunomodulatory Function of Mitochondrial and Nox-Derived Reactive Oxygen and Nitrogen Species.” Free Radical Biol and Med 109: 48-60.
- Jayaram S, Krishnamurthy PT (2021) Role of microgliosis, oxidative stress and associated neuroinflammation in the pathogenesis of Parkinson’s disease: The therapeutic role of Nrf2 activators. Neurochem Int 145: 105014.
- Quiñones Mar, RenéHernández-Bautista, DanielBeiroa, VioletaHeras, Francisco L, et al. (2021) “Sirt3 in POMC Neurons Controls Energy Balance in a Sex- and Diet-Dependent Manner.” Redox Biol 41: 101945.
- Denise D Belsham, Prasad S Dalvi (2020) Insulin signalling in hypothalamic neurones. J Neuroendocrinol 33: e12919.
- Huxing Cui, Miguel López, Kamal Rahmouni (2017) “The Cellular and Molecular Bases of Leptin and Ghrelin Resistance in Obesity.” Nature Reviews Endocrinol 13: 338-51.
- Ramadori Giorgio,Teppei Fujikawa, Makoto Fukuda, Jason Anderson, Donald A Morgan, et al. (2010) “SIRT1 Deacetylase in POMC Neurons Is Required for Homeostatic Defenses against Diet-Induced Obesity.” Cell Metabol 12: 78-87.
- Ritossa F (1962) “A New Puffing Pattern Induced by Temperature Shock and DNP in Drosophila.” Experientia 18: 571-3.
- J M Matz, MJ Blake, HM Tatelman, KP Lavoi, NJ Holbrook (1995) Characterization and regulation of cold-induced heat shock protein expression in mouse brown adipose tissue. Am J Physiol 269: R38-47.
- Y Cao, N Ohwatari, T Matsumoto, M Kosaka, A Ohtsuru, et al. (1999) TGF-beta1 mediates 70-kDa heat shock protein induction due to ultraviolet irradiation in human skin fibroblasts. Pflugers Arch 438: 239-44.
- Alain F Laplante, Véronique Moulin, FA Auger, J Landry, H Li, et al. (1998) Expression of Heat Shock Proteins in Mouse Skin During Wound Healing. J Histochem and Cytochem 46: 1-11.
- “Heat Shock Protein.” Wikipedia, 23 Apr. 2021, en.wikipedia. org/wiki/Heat_shock_protein#cite_note-pmid9921710-5. Accessed 1 May 2021.
- “Heat Shock Transcription Factors: Structure and Regulation.” Annual Reviews, 2019.
- Li, Zihai, Pramod Srivastava (2004) “Heat-Shock Proteins.” Current Protocols in Immunology.
- HA Parag, B Raboy, RG Kulka (1987) “Effect of Heat Shock on Protein Degradation in Mammalian Cells: Involvement of the Ubiquitin System.” The EMBO J 6: 55-61.
- NP Walsh, Benjamina MA, B Bose, CA Gross, Robert TS (2003) OMP Peptide Signals Initiate the Envelope-Stress Response by Activating DegS Protease via Relief of Inhibition Mediated by Its PDZ Domain. Cell 113: P61-71.
- Franz Narberhaus (2010) Translational control of bacterial heat shock and virulence genes by temperature-sensing mRNAs. RNA Biol. 7: 84-9.
- Petersen NS, HK Mitchell (1981) “Recovery of Protein Synthesis after Heat Shock: Prior Heat Treatment Affects the Ability of Cells to Translate MRNA.” Proceedings of the National Academy of Sci 78: 1708-11.
- HK Mitchell, P Young, NS Petersen (1988) “Heat Shock Protection against Cold Stress of Drosophila Melanogaster.” Mol Cell Biol 8: 3550-2.
- Walter, Stefan, Johannes Buchner (2002) “Molecular Chaperones—Cellular Machines for Protein Folding.” Angewandte Chemie International Edition 41: 1098-113.
- Julio C Borges, Carlos HI, Ramos (2005) “Protein Folding Assisted by Chaperones.” Protein & Peptide Letters, 12: 257-61.
- Mitchell HK, NS Petersen, CH Buzin (1985) Self-Degradation of Heat Shock Proteins.” Proceedings of the National Academy of Sci 15: 4969-73
- Benjamin, Ivor J, D Randy McMillan (1998) “Stress (Heat Shock) Proteins.” Circ Res 83: 117-32.
- Gabriel E Rosenfeld, Emily J Mercer, Christopher E Mason, Todd Evans (2012) Small heat shock proteins Hspb7 and Hspb12 regulate early steps of cardiac morphogenesis. Devel Biol 381: 389-400.
- Fan Guo-Chang, X Ren, J Qian, Q Yuan, Y Wang, et al. (2005) “Novel Cardioprotective Role of a Small Heat-Shock Protein, Hsp20, against Ischemia/Reperfusion Injury.” Circulation 111: 1792-9
- Deffit, Sarah N, Janice S Blum (2015) “A Central Role for HSC70 in Regulating Antigen Trafficking and MHC Class II Presentation.” Mol Immunol 68: 85-8.
- Ayesha Murshid, Jianlin Gong, Stuart K Calderwood (2012) “The Role of Heat Shock Proteins in Antigen Cross Presentation.” Frontiers in Immunol 3: 63.
Artcle Information
Review Article
Received Date: June 05, 2025
Accepted Date: June 23, 2025
Published Date: June 30, 2025
Journal of Neuroscience Research and Alzheimer’s Disease
Volume 1 | Issue 1
Citation
M Zubair Baba, Gowmathy S, Umair Wahedi (2025) Therapeutic Potential of NRF2 pathway in Neurological Disorder. J Neurosci Res Alzheimers Dis 1: 103
Copyright
©2025 M Zubair Baba. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

doi: jnra.2025.1.103