

Review Article
Volume-1 Issue-2, 2025
Worldwide status of insecticide resistance of Head louse (Pediculus humans capitis) (A Review)
Received Date: June 04, 2025
Accepted Date: June 23, 2025
Published Date: June 30, 2025
Journal Information
Abstract
Head lice infection is recognized as a major public health problem worldwide. Lice have shown resistance to insecticides, and as a result, these insecticides have lost their effectiveness. The main objective is to find the status of insecticide resistance to different insecticides in the world. An intensive search of scientific literature was reviewed using the search term in the following databases: “PubMed”, “Web of Knowledge”, “Scopus”, “Google Scholar”, “SID”, etc. The WHO guideline was considered for insecticide resistant level. There is now strong evidence of insecticide resistance in many countries to the point that some of these chemicals are obsolete. The lack of a local monitoring system for resistance patterns has made head lice treatment difficult. Clear treatment guidelines developed by health care professionals for cypress lice should follow local/national resistance patterns.The results is generalizable to real-world populations.These guidelines should combine chemical and non-chemical approaches to treatment and be reviewed by local public health departments in a coordinated and regular manner. Effective pharmaceutical companies must be present to deliver the up-to-date effects of their products.
Key words
Insecticide; Resistant, Pediculus humans capitis;world
 |
| Figure 1: The life cycle of head lice (Pediculosis capitis) and disease symptoms in humans |
 |
| Figure 2:Map of geographical distribution of pediculosis capitis worldwide(35) |
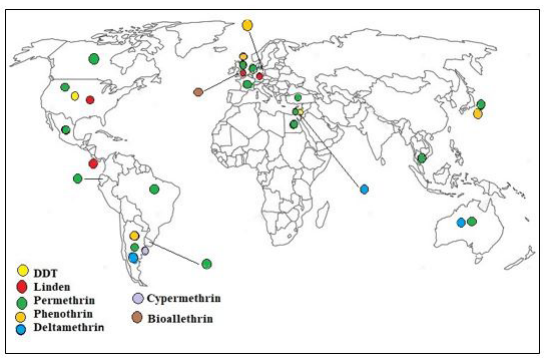 |
| Figure 3: The resistance of Pediculus humans capitis against organochlorine and Pyrethroids insecticides in different regions of world |
 |
| Figure 4: The resistance of Pediculus humans capitis against Organophosphates and Carbamates insecticides in different regions of world |
Introduction
Human head lice are blood-sucking insects that cause infections in humans called Pediculosis capitis. The P. capitis is a hematopoietic ectoparasite that is the most common parasitic infection in humans [1]. This infection is more common in the school-age population [2]. Itching of the scalp is a common symptom that causes itching and sleeps disturbance (Figure 1). These conditions can turn into a social fingertip that can lead to a loss of self-esteem [3]. Although P. humanus capitis is not a vector of human disease and poses no risk to human health, head lice infection can cause social distress, upset, parental anxiety, embarrassment for the child, and unnecessary absenteeism from school and work (Figure 2) [4]. The use of insecticides is the most effective method in the treatment of pediculosis. Pediculicides such as natural Pyrethrin (Pyrethrum),synthetic Pyrethroids (Permethrin, Phenothrin), Organochlorine (Indole), Organophosphorus (Malathion), and Carbamate (Carbaryl) are commonly used to treat lice. Chemical control of head lice is based on common insecticide species such as DDT and Lindane in the 1940s-1950s, Carbaryl and Malathion in the 1960s-1970s, and Pyrethroids in the 1980s. Frequent use of compounds in all these classes has caused resistance in the country [5]. Extensive use of insecticides and lack of proper replacement causes resistance to pediculosis. Resistance to topical fungicides is one of the emerging concerns in most parts of the world. One of the mechanisms of resistance to Pyrethrins or Pyrethroids is the insensitivity of the target area to impact resistance (kdr), a hereditary trait. Three-point mutations (M815I, T917I, and L920F) were found in the membrane segment, and this haplotype was present in the Permethrin -resistant head lice population [6]. With increasing treatment failure, issues such as misdiagnosis, non-adherence, inadequate treatment, re-infection, lack of ovarian or fungal remnant properties, and/or lice resistance to Pediculicides should be considered. The problem is directly related to the frequency used in the genotype and shows that head lice are resistant to similar DDT, Pyrethrin, and Pyrethroids [7]. Due to environmental concerns and fears of increasing lice resistance to Pediculicides, another factor was created to change the country's insects. Mutual resistance too many Pediculicides may contribute to a new wave of head lice infestation [8]. Mapping and understanding resistance mechanisms are essential for developing more effective treatments as well as extending the life of existing pedicures [9]. This inefficiency is mainly due to various reasons such as sales of ineffective products, misuse of Pediculicides, and the fact that lice in several countries develop resistance to insecticides such as DDT, Lindane, Malathion, Carbaryl, Permethrin, and δ-Phenothrin [10]. Resistance to head lice products has been demonstrated in several countries, including Canada, the United Kingdom, Korea, Japan, the United States, and Israel [11]. The high selection pressure caused by neuronal insecticides has led to the emergence and development of resistance to organochlorines, Pyrethroids, and organophosphorus compounds in many parts of the world. Indeed, clinical, parasitic, and molecular data have shown that conventional topical toxins worldwide have suffered significant loss of activity [12].
Methods
All published articles by search terms “resistance” and “pediculus humans capitis”, “Pyrethroid”, “Organochlorines”, “Organophosphate”, “Carbamate“, IGR”, and “kdr”, “P450”, “Monooxygenase”, “Glutathione”, or “Esterase” were conducted . Data were extracted from all articles. An intensive search of scientific literature was reviewed using the search term in the following databases: “PubMed”, “Web of Knowledge”, “Scopus”, “Google Scholar”, “SID”, etc. The WHO guideline was considered for insecticide resistant level.
Results and discussion
Resistant status of Pediculus humans capitis to Organochlorines insecticides
Resistance of the human head lice, P. capitis (Phthiraptera: Pediculidae), to Organochlorine insecticides such as DDT and Lindane has been recorded worldwide including Israel, Canada, Denmark, and Malaysia [9].
In America
The DDT resistance was demonstrated in the 1960s. Lindane resistance has been reported among head lice in the United States, United Kingdom, Netherlands, and Panama. Changes in amino acids located in the neural sheath sodium channel receptor have been described as the most important state of resistance and may cause cross-resistance to DDT, Lindane, and Pyrethroids (the oldest and most Permethrin)(8). In Panama, where the initial resistance to natural elders is minimal, the effect of treatment varies only from 30 (for R&C) to 52 (for RID) [8].
In Asia
DDT was the mainstay of lice control in Israel until the early 1980s when DDT resistance developed in local cap lice [13,14]. In Israel, the head lice, resisted DDT through the use of this insect until the 1980s [15]. Identification of the mechanism of DDT resistance based on glutathione S-transferase (GST) in Israeli head lice. This GST mechanism occurred before 1989, while Permethrin resistance was established in P. capitis after 1994, indicating that the main mechanism of GST selection using DDT does not provide any Pyrethroids cross-resistance [15].
Resistant status of P. humans capitis to Pyrethroids
Pyrethroids become excellent insecticides due to their low mammalian toxicity and relatively harmless environmental residues. However, resistance to this type of insect has been well documented in the human head lice population of the country, including the United Kingdom, France, Japan, Israel, the United States, Argentina and Denmark, Mexico, Russia and Chile (Figure 3) [16,17]. Recently, resistance to Permethrin has been widely reported in several countries, including France (1993), the Czech Republic (1995), the Middle East, Israel (1995), the United Kingdom (1995, 1999), and South America (Argentina). (1998, 2000), and North America (USA, 1999). In Europe (UK and Denmark), Asia (Japan) and Australia .There are several mechanisms of Permethrin resistance, the most important of which is through the kdr gene [8].
In Asia
In Israel, significant changes in Permethrin susceptibility were observed in head lice collected in 1989 and 1994 [14]. Pyrethroids resistance has developed rapidly among head lice since the introduction of Permethrin as a pedicure in 1991 [13]. Therefore, it seems probable that Pyrethroids resistance in Israeli is due to a combination of neurosensitivity (shock resistance or "kdr") and monooxygenase resistance mechanisms [15]. The rapid development of Permethrin is due to cross-resistance of DDT due to impact resistance (kdr) [13]. Mechanism examination of lice from a large number of infected people in a large city such as Jerusalem shows that resistance to Permethrin is very widespread in Israel, not only due to the high mutation rate and high percentage of resistant lice but also, none of the resistant lice were resistant to only one allele [14]. The Pyrethroids resistance of in Israel extends to Phenytoin [15]. The first study to detect Permethrin resistance among human head lice from Thailand showed that the nucleotide and amino acid sequences of RS and RR show the T917I and L920F point mutations. The data from this study raise awareness of the increasing kdr mutation in head lice in Thailand. The head lice kdr mutation in Thailand showed three different genotypes [3]. The first large-scale Pyrethroids-resistant head lice in Japan showed that colony-resistant genes were collected from 11 of 22 provinces and it was speculated that power lice are currently being developed specifically in Japan [11]. High resistance to Phenothrin was recorded for Japan. In Okinawa Prefecture, 96% of head lice are resistant to Pyrethroids [18]. The results of a study screened for Permethrin resistance in the Turkish head lice population showed the frequency of high-strength alleles among the samples [19].
In America
Pyrethroids resistance leads to disruption of treatment and the development of chronic infections, which cost approximately $ 1 billion a year in the United States alone [20]. Permethrin was first used in Argentina in 1990, and resistance to Permethrin was first diagnosed in 1997 in various populations in Buenos Aires [21]. In a 2003 study in different parts of Buenos Aires, where the paper exposure method had previously recorded high levels of Permethrin resistance [20]. An extensive survey on resistance in Buenos Aires found that there was a high level of head lice in children in 24 of 26 schools (92.3%) [21]. All lice resistant to Permethrin (resistance ratio 52.8 to 0.88.7) were also resistant to d- Phenothrin (resistance ratio 40.86 to 0.48.39) and Deltamethrin (resistance ratio 16.24 to 38.06). Otherwise, all colonies resistant to low to high levels showed resistance to b-cypermethrin. This Pyrethroids has never been used as a Pediculicides in Argentina [5]. However, the high level of resistance in these Permethrin-resistant colonies (resistance ratio 9.74 to 50.9). The high resistance of Pyrethroids in Buenos Aires can be due to the extreme selection of Permethrin. In addition, because b-cypermethrin has never been used as a Pediculicides in Argentina, high levels of resistance in Permethrin-resistant colonies indicate that there is pyrethyroid resistance [22] with 30 years of use of Pyrethroids-based Pediculicides. In Argentina, kdr resistance has evolved rapidly among these lice population [16]. In different parts of Buenos Aires, lice colonies with Permethrin resistance showed resistance to Deltamethrin 1998. This is the first evidence that head lice in Chile have mutations that are commonly associated with Pyrethroids resistance [22]. Insect-resistant populations were obtained from Florida (FL-HL) and Massachusetts (MAHL) in the United States and Bristol (BR-HL) in the United Kingdom [23]. The presence of T929I and L932F in Permethrin-resistant South Florida lice was confirmed by DNA sequencing. The loss of performance of the Nix formula (Prestige Brand, Tarrytown, NY) from 1998 to 2013 was associated with an increase in kdr-type mutations. These data provide a plausible reason for the reduced effect of Permethrin in the Nix formula, which is a parallel increase in kdr-type mutations in lice over time [24]. In Uruguay and Brazil, Pyrethroids resistance in kdr, and in 2017, in Mexico showed Pyrethroids resistance due to kdr [9]. The presence of a C / T mutation responsible for the T917I kdr allele associated with Pyrethroid resistance in P. capitis from Honduras(17). Biomarker of kdr- Permethrin resistance was detected for T917I mutation by RFLP and PCR in 99.9% of head lice tested from Georgia(25). The prevalence of resistant allele (R) in head lice from Canada could indicate failure of treatment with pyrethrin and Pyrethroids. [26] The representative demographic sample was tested using biochemical and molecular methods and it was shown that in Wales, treatments containing Pyrethroids were less effective in controlling head lice infection than those containing organophosphate [27].
In Africa
In Egypt in 2010, Clark's studies showed Pyrethroids resistance in the region [9].
In Europe
The presence of Pyrethroid-resistant head lice for more than 20 years has been reported mainly in Europe [18]. In the UK, a 1998 local survey of 10 schools in Bristol and Bath showed resistance to treatment with Malathion and Permethrin in head lice collected from infected school children [21]. A similar situation has been reported by Burgess and Brown (1999) from several regions in the United Kingdom. The cap from the Brighton area, on the south east coast of England, showed 400-fold resistance to Malathion, and resistance to Permethrin occurred at a lower level than Malathion. A similar pattern of Malathion and Permethrin appeared in other parts of the United Kingdom [21]. In 2000 the study in the United Kingdom found that there was high resistance to Permethrin, Phenothrin, and Malathion with 87% Permethrin failure rate and 64% Malathion failure rate with topical treatment [4,17]. Permethrin resistance in Czechoslovakia head lice was associated with cross-resistance to D-Phenothrin and Bioallethrin, and French lice were also resistant to D-Phenothrin. In 1998, one study reported that all desert lice that were resistant to Permethrin also became resistant to D-Phenothrin and Deltamethrin. Therefore, most Permethrin-resistant lice were resistant to other Pyrethroids insecticides. High levels of resistance to D-Phenothrin, Deltamethrin, and b-Permethrin were observed [22]. Resistance to several Pyrethroids has been reported by Rups (1995). In the head lice population of the Czech Republic. The authors reported that P. capitis is resistant to Permethrin and that Pyrethroids cross-resistance includes Bioallethrin and Diphenothrin(22). In Australia and Denmark, Clark showed Pyrethroids resistance in kdr, as well as in Ecuador, NA resistance(9). The prevalence of Permethrin and Malathion resistant head lice is high in the Danish head lice population(28). The association between Permethrin resistance and kdr-like mutations is confirmed by our findings. The frequency of T929I-L932 F dual mutations in the voltage-sensitive sodium channel gene was associated with 0.95 Permethrin resistance in the Danish head lice population [28]. The presence of Pyrethroids-resistant genes in the head lice population of students in an urban area in France was studied [1].
In Australia and Oceania
While head lice are resistant to several Pyrethroids (including natural pyrethrum and synthetic Permethrin), Malathion and its associated compounds have been confirmed abroad and have been suspected in Australia, 5 treatments in Australia are still due to misdiagnosis poor use of pediculicides or re-accumulation [29] (Figure 3).
Resistant status of Pediculus humans capitis to Organophosphates insecticides
In Europe:In the UK, a local survey of 10 schools in Bristol and Bath in 1998 found resistance to Malathion treatment for head lice collected from school children. In addition, high resistance to Malathion was measured for head lice from the UK(30). Also Malathion resistance was reported in the UK in 1999 [31].
Resistant status of Pediculus humans capitis to Carbamates insecticides
The effect of Carbaryl Shampoo on head lice has decreased due to its worldwide resistance. Carbaryl shampoo resistance of head lice has been reported in several countries such as Australia, the United Kingdom, and the United States [32].
In America
Because there is resistance to head lice in different parts of the UK against all over-the-counter products containing synthetic insecticides, and resistance to Carbaryl appears to be emerging, the adult head lice found on Leeds children after treatment, was probably resistant to Carbaryl [30,33] (Figure 4).
Mechanisms of insecticide resistance in Pediculus humans capitis
Most of the resistance mechanisms studied is through Knockdown resistance mutations (kdr) with biomarkers M815I, T917I and L920F, which are mostly reported in lice. Other mechanisms such as cytochrome P450 inhibition, acetylcholinesterase binding, and GluCl mutations have been reported(9). The results presented in the case of cross-resistance and synergistic proliferation by enzyme inhibitors in Permethrinresistant head lice showed that increased metabolism is involved in Pyrethroids resistance. However, the significant amount of resistance that remained after synergy indicated the existence of another resistance mechanism. Pyrethroids cross-resistance and Carbaryl carbamate sensitivity suggested a common mechanism of action [22]. Allelic mutations associated with Pyrethroid resistance vary in regions and countries, indicating the need to monitor resistance through region-specific genetic markers. In well-developed geographical areas, the lice population is predominantly homozygous. In areas where heterozygosity is still present, selective pressure using chemical bases increases resistance [31].
Conclusions
Currently, the recommended therapies include Pyrethroids, which are known as the first line of treatment. But resistance to these neurotransmitters has become increasingly common. Alternative therapies such as newer neuroleptics or Pediculicides with physical mechanisms of action (e.g., dimethicone-based products) should be considered, especially in areas with high resistance. Continuous resistance monitoring is required to monitor the frequency of resistance alleles, which is expected to increase to a fixed state as the lice population increases. Finally, Head lice training, proper use of treatment, and coordinated diagnosis are essential for effective management of this problem. More research needs to be done on new therapies with a mechanism of physical activity. World Health Organization(34) recommended the following insecticide for lice control Carbaryl, Propoxur, Lindane, Malathion, Temephos, Bioallethrin, Permethrin, Diphenothrin. Monitoring of resistance of head lice to these insecticides is an important factor for decision-makers.
References
- Durand R, Millard B, Bouges-Michel C, Bruel C, Bouvresse S, Izri A. 2007. Detection of pyrethroid resistance gene in head lice in schoolchildren from Bobigny, France. J. Med. Entomol; 44(5):796-8.
- Eremeeva M, Anderson M, Ogunleke T, Warang S, Capps D, Zohdy S, et al. 2018. Pathogens, pesticide resistance and genetic diversity of human head lice. Zoonotic . Parasitic Infect; 8(4):545-549.
- Brownell N, Sunantaraporn S, Phadungsaksawasdi K, Seatamanoch N, Kongdachalert S, Phumee A, et al. 2020. Presence of the knockdown resistance (kdr) mutations in the head lice (Pediculus humanus capitis) collected from primary school children of Thailand. PLOS Negl. Trop. Dis; 14(12):e0008955.
- Bharti S, Duragkar N.2017. A review on Pediculus humanus capitis: Based on life cycle, resistance, safety considerations and treatment. Int. J. Indig. Herbs. Drugs. (4)1:27-36.
- Hatam-Nahavandi K, Ahmadpour E, Pashazadeh F, Dezhkam A, Zarean M, Rafiei-Sefiddashti R, et al. 2020. Pediculosis capitis among school-age students worldwide as an emerging public health concern: a systematic review and meta-analysis of past five decades. Parasitol. Res; 1-19.
- Toloza AC, Zygadlo J, Cueto Gm, Biurrun F, Zerba E, Picollo MI. 2006. Fumigant and repellent properties of essential oils and component compounds against permethrin-resistant Pediculus humanus capitis (Anoplura: Pediculidae) from Argentina. J. Med. Entomol. 2006;43(5):889-95.
- Ertabaklar H, Malatyali E, Ertug S. 2020. Drug resistance in parasitic diseases. Eur. J; 26:1-5.
- Husni L, Al-Waiz M. 2021. Topical ivermectin in the treatment of Pediculosis capitis. Our Dermatology Online/Nasza Dermatol Online.;12(1).16-23.
- Jones KN, English JC. 2003. Review of common therapeutic options in the United States for the treatment of pediculosis capitis. Clin. Infect. Dis; 36(11):1355-61.
- Fox K, Larkin K, Sanchez A.2020. Global trends in genetic markers of pediculus humanus capitis resistance mechanisms. Curr. Trop. Med . Rep;7:65-73.
- Toloza AC, Lucía A, Zerba E, Masuh H, Picollo MI. 2010. Eucalyptus essential oil toxicity against permethrin-resistant Pediculus humanus capitis (Phthiraptera: Pediculidae). Parasitol. Res;106(2):409-14.
- Kasai S, Ishii N, Natsuaki M, Fukutomi H, Komagata O, Kobayashi M, et al. 2009. Prevalence of kdr-like mutations associated with pyrethroid resistance in human head louse populations in Japan. J. Med. Entomol ;46(1):77-82.
- Candy K, Nicolas P, Andriantsoanirina V, Izri A, Durand R. 2018. In vitro efficacy of five essential oils against Pediculus humanus capitis. Parasitol. Res;117(2):603-9.
- Mumcuoglu KY, Hemingway J, Miller J, IOffe‐uspensky I, Klaus S, Ben‐ishaI F, et al. 1995. Permethrin resistance in the head louse Pediculus capitis from Israel. Med. Vet. Entomol;9(4):427-32.
- Lindh J, Mumcuoglu KY. 2012. Molecular analysis of pyrethroid resistance in Pediculus humanus capitis from Israel. Trends. Entomol;8:37-41.
- Hemingway J, Miller J, Mumcuoglu K. 1999. Pyrethroid resistance mechanisms in the head louse Pediculus capitis from Israel: implications for control. Med. Vet. Entomol;13(1):89-96.
- Roca-Acevedo G, del Solar Kupfer CP, Dressel Roa P, Toloza AC.2019. First determination of pyrethroid knockdown resistance alleles in human head lice (Phthiraptera: Pediculidae) from Chile. J. Med. Entomol ;56(6):1698-703.
- Larkin K, Rodriguez CA, Jamani S, Fronza G, Roca-Acevedo G, Sanchez A, et al. 2020. First evidence of the mutations associated with pyrethroid resistance in head lice (Phthiraptera: Pediculidae) from Honduras. Parasit Vectors. 13(1):1-7.
- Yamaguchi S, Yasumura R, Okamoto Y, Okubo Y, MiyagiT, Kawada H, et al.2021. Efficacy and safety of a dimethicone lotion in patients with pyrethroid‐resistant head lice in an epidemic area, Okinawa, Japan. J . Dermatol. ; 48(9):1343-1349.doi: 10.1111/1346-8138.15966.
- Karakuş M, Atıcı T, Karabela ŞN, Baylan O, Limoncu ME, Balcıoğlu İC. 2020. Detection of permethrin resistance and phylogenetic clustering of turkish head lice (Pediculus humanus capitis; De Geer, 1767 populations. Acta .Trop ;204:105362.
- Clark JM. 2009. Determination, mechanism and monitoring of knockdown resistance in permethrin-resistant human head lice, Pediculus humanus capitis. J. Asia-Pacific .Entomol ;12(1):1-7.
- Vassena CV, Cueto GM, Audino PG, Alzogaray RA, Zerba EN, Picollo MI. 2003. Prevalence and levels of permethrin resistance in Pediculus humanus capitis De Geer (Anoplura: Pediculidae) from Buenos Aires, Argentina. J. Med. Entomol ;40(4):447- 50.
- Picollo M, Vassena C, Casadio A, Massimo J, Zerba E. 1998. Laboratory studies of susceptibility and resistance to insecticides in Pediculus capitis (Anoplura; Pediculidae). J. Med. Entomol ;35(5):814-7.
- Lee SH, Yoon K-S, Williamson MS, Goodson SJ, Takano-Lee M, Edman JD, et al.2000. Molecular analysis of kdr-like resistance in permethrin-resistant strains of head lice, Pediculus capitis. Pest. Biochem .Physiol; 66(2):130-43.
- Gellatly KJ, Krim S, Palenchar DJ, Shepherd K, Yoon KS, Rhodes CJ, et al. 2016. Expansion of the knockdown resistance frequency map for human head lice (Phthiraptera: Pediculidae) in the United States using quantitative sequencing. J. Med. Entomol ;53(3):653-9.
- Eremeeva ME, Capps D, Winful EB, Warang SS, Braswell SE, Tokarevich NK, et al. Molecular markers of pesticide resistance and pathogens in human head lice (Phthiraptera: Pediculidae) from rural Georgia, USA. J. Med. Entomol;54(4):1067-72.
- Marcoux D, Palma KG, Kaul N, Hodgdon H, Van Geest A, Previte DJ, et al. 2010. Pyrethroid pediculicide resistance of head lice in Canada evaluated by serial invasive signal amplification reaction. J. Cutan. Med. Surg;14(3):115-8.
- Thomas DR, McCarroll L, Roberts R, Karunaratne P, Roberts C, Casey D, et al. 2006. Surveillance of insecticide resistance in head lice using biochemical and molecular methods. Arch. Dis. Child ;91(9):777-8.
- Kristensen M, Knorr M, Rasmussen A-M, Jespersen JB. 2006. Survey of permethrin and malathion resistance in human head lice populations from Denmark. J. Med. Entomol.;43(3):533-8.
- Bailey AM, Prociv P.2004. Persistent head lice following multiple treatments: evidence for insecticide resistance in Pediculus humanus capitis. Aust. J .Dermatol ;41(4):250-4.
- Audino PG, Vassena C, Zerba E, Picollo M2007. Effectiveness of lotions based on essential oils from aromatic plants against permethrin resistant Pediculus humanus capitis. Arch. Derm. Res;299(8):389-92.
- Yoon KS, Gao J-R, Lee SH, Clark JM, Brown L, Taplin D. 2003. Permethrin-resistant human head lice, Pediculus capitis, and their treatment. Arch. Dermatol ;139(8):994-1000.
- Sittichok S, Soonwera M. 2018. Efficacy of New Herbal Shampoos from Garcinia dulcis Kurz, Citrus aurantium L. and Eucalyptus globulus Labill as Pediculicides for Head Lice (Pediculus humans capitis) Control. Inter. J .Agri. Technol;14(4):597-612.
- Downs A, Stafford K, Hunt L, Ravenscroft J, Coles G. 2000. Widespread insecticide resistance in head lice to the over‐the‐ counter pediculocides in England, and the emergence of carbaryl resistance: Therapeutics. Br. J. Dermatol; 146(1):88-93.
- World Health Organization (WHO). 2006. Pesticides and their application: for the control of vectors and pests of public health importance. World Health Organization, 2.pp.151.
Artcle Information
Review Article
Received Date: June 04, 2025
Accepted Date: June 23, 2025
Published Date: June 30, 2025
Annals of Public Health and Preservative Medicine
Volume 1 | Issue 2
Citation
Ghazal Tashakori (2025) Worldwide status of insecticide resistance of Head louse (Pediculus humans capitis) (A Review). Ann Public Health Preservative Med 1: 202
Copyright
©2025 Hassan Vatandoost. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.

doi: appm.2025.1.202